










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkArchivos de Zootecnia
versión On-line ISSN 1885-4494versión impresa ISSN 0004-0592
Arch. zootec. vol.59 no.227 Córdoba sep. 2010
Doses de fósforo e inoculação micorrízica no cultivo de estilosantes em solo sob condições naturais*
Phosphorus levels and mycorrhizal inoculation growing stylosanthes on the soil under natural conditions
Carneiro, R.F.V.1, Martins, M.A.2, Vásquez, H.M.3 e Detmann, E.4
1UFPI. Campus Bom Jesus/PI. Brasil. romero@ufpi.edu.br
2UENF/CCTA/Laboratório de Solos. Brasil. marco@uenf.br
3UENF/CCTA/Laboratório de Zootecnia. Brasil. maldonado@uenf.br
4Departamento de Zootecnia/UFV. Viçosa, MG. Brasil. detmann@ufv.br
*Projeto financiado pela FAPERJ executado na Universidade Estadual do Norte Fluminense Darcy Ribeiro-UENF.
RESUMO
Este trabalho conduzido em casa de vegetação teve por objetivo avaliar a resposta do estilosantes à inoculação com fungos micorrízicos arbusculares e aplicação de doses de fósforo em solo não esterilizado (condições naturais). Foi utilizado um Latossolo Amarelo distrófico. O delineamento experimental foi em blocos casualizados, num esquema fatorial 4 x 3, sendo quatro doses de P (0, 60, 120 e 240 mg/dm3 de solo) e três tratamentos microbiológicos (controle; inoculação com o fungo Glomus clarum e inoculação com o inóculo nativo), com 3 repetições. Realizou-se dois cortes da parte aérea, a cada 60 dias. Analisou-se a produção de matéria seca da parte aérea e raiz, os acúmulos de proteína bruta, P, K, Ca, Mg e S; percentagem de colonização micorrízica e densidade de esporos. Os resultados demonstraram que o aumento das doses de P incrementaram significativamente as variáveis estudadas. Esses resultados foram evidenciados pela presença da inoculação micorrízica, com destaque para a presença da espécie Glomus clarum, principalmente para o primeiro corte. Estimou-se uma redução de 82,8 e 71,3% no requerimento externo de P, para o estabelecimento do estilosantes em função da inoculação com o Glomus clarum e inóculo nativo, respectivamente. No segundo corte, verificou-se efeito dos tratamentos micro-biológicos, somente para as quantidades acumuladas de Ca e Mg. A colonização micorrízica e a densidade de esporos não foram afetadas pelos tratamentos microbiológicos.
Palavras chave: Forrageira. Glomus clarum. Inóculo nativo. Nutrição mineral. Solo não esterilizado.
SUMMARY
This work, carried out under greenhouse, had as objective to evaluate the response of the Stylosanthes to mycorrhizal inoculation and levels of phosphorus in soil no sterilized (natural conditions). A randomized blocks design was used in a 4 x 3 factorial scheme (four phosphorus levels: 0, 60, 120 and 240 mg/dm3 of soil; three microbiology treatments: control; inoculation with Glomus clarum and inoculation with native inoculum), with three repetitions. Two cuts of the aerial part were accomplished every 60 days. It was analyzed the production of dry matter of the aerial part and root, the accumulations of crude protein, P, K, Ca, Mg and S, percentage of mycorrhizal colonization and density of spores. The results demonstrated that the increase of the doses of P increased significantly the studied variables. Those results were evidenced by the presence of the mycorrhizal inoculation, with prominence for the presence of the species Glomus clarum, mainly for the first cut. Was estimated a reduction of 82.8 and 71.3% in the external requirement of P, for the establishment of the Stylosanthes in function of the inoculation with the Glomus clarum and native inoculum, respectively. In the second cut, effect of the microbiology treatments was verified, only for the accumulated amounts of Ca and Mg. The mycorrhizal colonization and the density of spores were not affected for the microbiology treatments.
Key words: Forage. Glomus clarum. Native inoculum. Mineral nutrition. No sterilized soil.
Introdução
A região Norte/Noroeste Fluminense, Estado do Rio de Janeiro-Brasil, é caracterizada por solos ácidos e de baixa fertilidade natural, com evidente processo de degradação pelo monocultivo da cana-de-açúcar, e são constituídos basicamente por argilas de baixa atividade (Fontes, 2001). Em levantamento realizado por Tebaldi et al. (2000) em áreas ocupadas por pastagens nesta região, ficou constatado a deficiência generalizada principalmente de P e Ca, fato que pode contribuir para desencadear o processo de degradação das pastagens e comprometer a viabilidade de introdução das leguminosas forrageiras no sistema produtivo.
As reduções em produtividade e valor nutricional das plantas forrageiras ocorrem em razão do manejo inadequado dos solos tropicais, acarretando em reduções em fertilidade. Fato que compromete, principalmente, a introdução e estabelecimento das leguminosas forrageiras (Oliveira et al., 1996). No entanto, o estilosantes é uma leguminosa forrageira que tem apresentado excelente adaptação a solos ácidos e de baixa fertilidade, eficiente fixação de N2 nestas condições e altos teores de proteína bruta na matéria seca (Andrade e Karia, 2000). Souza et al. (2000) descreveram o estilosantes como uma espécie altamente eficiente no uso de P, principalmente pela alta dependência à associação micorrízica.
As micorrizas arbusculares são asso-ciações simbióticas mutualistas entre raízes de plantas e fungos do filo Glomeromycota (Schussler et al., 2001), e favorecem o crescimento e aquisição de nutrientes pela grande maioria das plantas de interesse econômico. Nas últimas décadas, sobretudo pelo reconhecimento da importância funcional e ecológica da simbiose nos ecossistemas onde elas ocorrem, a importância dos fungos micorrízicos arbusculares (FMA) tornou-se mais evidente (Moreira e Siqueira, 2006).
Estes autores apontaram algumas questões recorrentes em micorrizologia, tais como: desenvolver tecnologias para a inoculação e/ou manejo das populações indígenas de FMA, e, estabelecer procedimentos eficazes para maximizar o potencial dos FMAs em programas de recuperação de áreas degradadas de baixa produtividade.
Destaca-se o manejo da simbiose micorrí-zica, como uma estratégia cujo propósito é a melhoria na nutrição e produtividade das pastagens com gasto mínimo de insumos, constiuindo-se em uma prática promissora para uma maior viabilidade e sustentabi-lidade de pastagens em solos tropicais.
Este estudo teve como objetivo avaliar a influência da inoculação de fungos micorrízicos arbusculares e da adubação fosfatada, sobre a produção e acúmulo de nutrientes na forragem do estilosantes, em solo sob condições naturais.
Material e métodos
O experimento foi conduzido em casa de vegetação do Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense (UENF), no período de novembro de 2002 a fevereiro de 2003. Utilizou-se uma amostra da camada superficial (0-20 cm) de um Latossolo Amarelo Distrófico Típico (LVd), sob pastagem nativa de capim pernambuco (Paspalum sp.). Após a coleta, a amostra do solo foi peneirada em malha de 2,0 mm e seco ao ar. Uma sub-amostra foi tomada para análises físico-químicas. Os resultados foram: pH em água - 5,3; P (Mehlich 1) 4,0 mg/dm3; K - 60,0 mg/dm3; Ca, Mg, Al, H+Al e Na - 1,6, 0,6, 0,2, 3,5 e 0,03 cmolc/dm3, respectivamente; matéria orgânica - 17,9 g/dm3; argila, silte e areia - 32, 8 e 60%, respectivamente, e a densidade média de esporos de FMA (a partir de 20 amostras ao acaso após peneiramento) foi de 24 esporos em 65 g de solo. Em seguida, a amostra foi submetida à calagem, pelo método da saturação por bases para se elevar o valor V para 60%, utilizando-se o calcário magne-siano comercial, com PRNT = 90%, e incubado por 20 dias em sacos plásticos.
O delineamento experimental utilizado foi em blocos casualizados, arranjado em esquema fatorial 4 x 3, com 3 repetições, sendo os fatores: quatro doses de P (0, 60, 120, e 240 mg/dm3) e três condições de solo (controle não inoculado-condição micro-biológica natural, solo inoculado com o FMA-Glomus clarum e solo reinoculado com os FMAs-nativos). Com essas dosagens utilizadas, objetivou-se garantir disponibilidades de P que variassem de baixo a alto.
Além das doses de P, foi aplicada uma adubação básica de semeadura, segundo Souza et al. (2000), que constou de 30 mg de N, 100 mg de K, 0,5 mg de B, 1,5 mg de Cu, 3,0 mg de Mn, 5,0 mg de Zn e 0,1 mg de Mo por dm3 de solo. As fontes utilizadas no preparo das soluções para adubação das parcelas foram os reagentes p.a.: NH4SO4, KH2PO4, NaH2PO4, K2SO4, KCl, H3PO4, H3BO3, CuCl2, MnCl2.4H2O, ZnSO4.7H2O, H2MoO4.H2O.
Os inóculos foram multiplicados em substrato solo:areia (proporção 2:1), autoclavado a 120oC por duas horas, tendo a Brachiaria brizantha como espécie hospedeira em vasos de três litros. O inóculo nativo foi obtido a partir da aplicação de 50 ml por vaso, do solo destinado a condução do experimento, misturado na superfície do vaso com o substrato solo:areia autoclavado. Posteriormente, a braquiária foi semeada e cultivada por 90 dias. O mesmo procedimento foi adotado para obtenção do inoculo Glomus clarum, no entanto, a alíquota de 50 ml por vaso, foi retida de câmara fria da coleção de FMA do laboratório de Micro-biologia do Solo da UENF. Após obtenção dos inóculos, o procedimento de inoculação foi feito no momento da semeadura aplicando-se a 5,0 cm de profundidade e abaixo das sementes, 300 ml (5% do volume do vaso de 6 l) de substrato-inóculo contendo esporos, raízes infectadas e pedaços de hifas.
O inóculo, da espécie micorrízica Glomus clarum, foi escolhido por sua efetividade demonstrada por trabalhos conduzidos na região (Schiavo e Martins, 2002; Rodrigues et al., 2003). O tratamento controle, recebeu 300 ml/vaso do mesmo substrato-inóculo, esterilizado em autoclave a 120oC por duas h.
A contagem de esporos de FMA no substrato-inóculo para os tratamentos Glomus clarum e inóculo nativo, foi de 488 e 298 em 65 g, respectivamente.
Posteriormente à emergência das plântulas do estilosantes, foram preparadas soluções com o substrato inóculo contendo a espécie Glomus clarum (solução A) e com o solo contendo o inóculo nativo (solução B), obtidas por suspensão de 10 cm3 de solo em seis litros de água, seguida por tamização em peneiras com aberturas de 0,710 e 0,053 mm e filtragem em papel de filtro para eliminação de propágulos, com objetivo de equilibrar a microbiota entre os tratamentos. As soluções obtidas, foram aplicadas para todas as parcelas do experimento.
A semeadura foi efetuada utilizando-se 20 sementes do estilosantes (Stylosanthes guianensis (Aubl.) cv. Mineirão) por vaso, deixando-se 4 plantas posteriormente. A umidade do solo foi mantida através de pesagens diárias dos vasos, amostrando-se aleatoriamente um vaso de cada dose de P, definindo-se a partir daí as quantidades de água em base de peso, a serem repostas por diferentes doses de P. Esse controle da umidade foi realizado baseando-se na manutenção de água em 60% do volume total de poros (VTP), definidos pela equação:
VTP=1 - Ds/Dp
onde:
Ds= densidade do solo
Dp= densidade de partículas
Os cortes foram realizados a 10 cm do solo, em intervalos de 60 dias o que coincidia (nas condições de casa de vegetação) com início de surgimento das primeiras folhas senescentes. O material colhido foi acondicionado em sacos de papel, previamente identificados. Esse material foi seco em estufa com circulação de ar a 65-70oC por 72 horas para obtenção da matéria seca. Posteriormente, esse material foi moído e armazenado em frascos devidamente etiquetados para determinação do rendimento de proteína bruta pelo método de Kjeldahl, e dos teores de P, K, Ca, Mg e S segundo metodologia descrita por Malavolta et al. (1989).
Foram retiradas amostras de radículas (nas porções superiores, medianas e terminais do sistema radicular) e colocadas no conservante álcool 50% para posterior avaliação da colonização micorrízica. No preparo da amostra para esta avaliação foi utilizado o método descrito por Giovannetti e Mosse (1980).
Para a quantificação de esporos no solo, as amostras compostas de 65 g de solo foram submetidas ao método de decantação e peneiramento úmido, segundo Gerdemann e Nicolson (1963), seguida de centrifugação em água por três minutos e em sacarose 50% por dois minutos. No material obtido, foi feita a contagem de esporos, com auxílio de microscópio.
Após a realização da análise de variância, procedeu-se o ajustamento de equações de regressão linear em superfície de resposta, incluindo-se o fator qualitativo tratamentos microbiológicos por intermédio de variáveis Dummy (Draper e Smith, 1996), adotando-se o modelo básico:
Yi= b0 + b1D1 + b2D2 + b3Pi + b4Pi2 + b5D1Pi + b6D2Pi + eij
em que:
b0= intercepto,
bj= coeficientes de regressão, sendo j= 1, 2, 3, 4, 5, 6,
D1 e D2= variáveis Dummy para o ajustamento com o fator categórico tratamento microbiológico, sendo D1= 0 e D2= 0 para o tratamento microbiológico controle; D1= 1 e D2= 0 para o tratamento microbiológico Glomus clarum, D1= 0 e D2= 1 para o tratamento microbiológico inóculo nativo,
Pi= dose de adubação fosfatada,
eij= erro aleatório, associado a cada observação, pressuposto NID (0, s2).
Resultados e discussão
MATÉRIA SECA DA PARTE AÉREA E SISTEMA RADICULAR
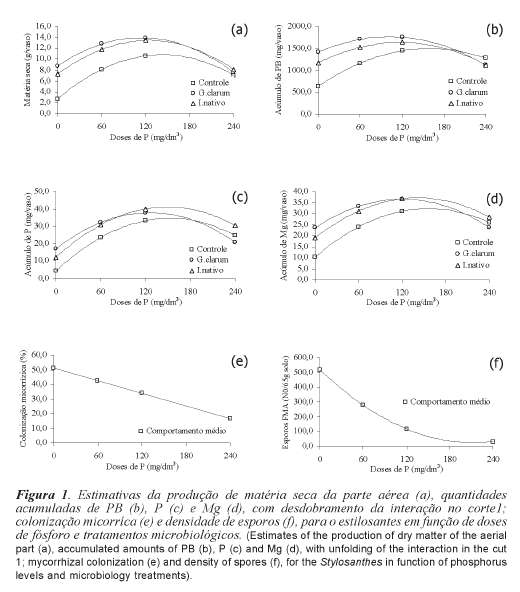
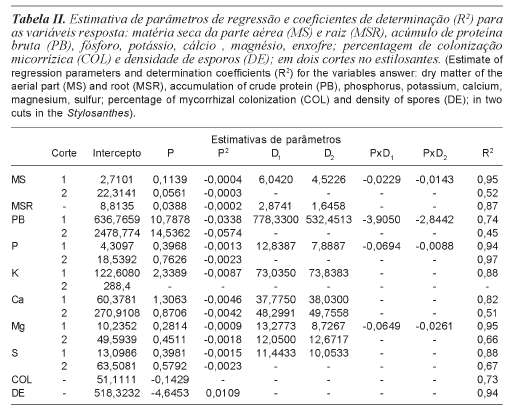
Para a produção de matéria seca da parte aérea (MS), no primeiro corte, verificou-se interação significativa entre P x tratamento microbiológico (p<0,05) (tabela I). Estudan-do o efeito das doses de P para os tratamentos microbiológicos, observa-se que os tratamentos de inoculação sobressaíram-se em relação ao controle principalmente nas doses inferiores aplicadas. Os benefícios da inoculação tenderam a diminuir com o aumento das doses (figura 1a), fato que corrobora com os resultados de Paulino et al. (1992), Santos et al. (2002), Carneiro et al. (1999) e Carneiro et al. (2002), que avaliaram o efeito da inoculação micorrízica e de doses de P em leguminosas forrageiras. O comportamento das doses foi descrito pelo modelo quadrático em todos os tratamentos microbiológicos (tabela II).
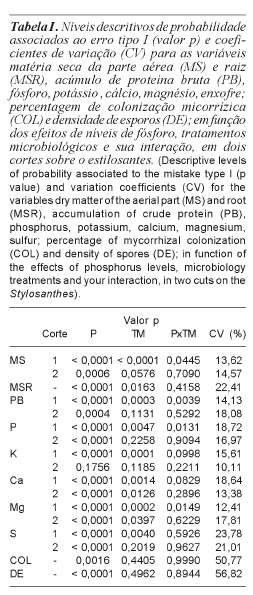
Para a produção de 10,8 g/vaso, correspondente à produção máxima obtida no tratamento microbiológico controle, necessitou-se da aplicação de 151,1; 26,0 e 43,3 mg/dm3 de P; para os tratamentos controle, Glomus clarum e inóculo nativo, respectivamente. Isso representou uma redução das necessidades de P para o estabelecimento da planta, da ordem de 82,8% para a inoculação com o Glomus clarum e 71,3% para a inoculação com o inóculo nativo. Reduções estas que mantiveram as mesmas proporções elevadas encontradas por Souza et al. (2000) estudando o estilosantes em substrato estéril.
Faria et al. (1995) concluíram que a magnitude das resposta a FMA por leguminosas está associada a fertilização com N e P associados a inoculação com rizóbio. Estes autores avaliaram FMA, nitrogênio e fósforo; em solo fumigado e não fumigado, e verificaram que a inoculação com FMA no solo não fumigado não promoveu crescimento à Albizia lebbeck (leguminosa arbórea) na ausência de fertilização com N e P. Entretanto, quando estes nutrientes foram aplicados em conjunto e também se adicionou rizóbio, a inoculação com FMA trouxe efeito significativo para o crescimento e aquisição de nutrientes. O solo apresentava uma contagem de 21 esporos de FMA nativos por 40 ml de solo, num período de 110 dias para a obtenção da muda, ou seja, condição semelhante ao presente estudo que teve duração de 120 dias.
Ying Chu et al. (2001) relacionaram ao fungo adaptado às condições edafoclimáticas a eficiência da prática da inoculação. Assim verificaram que em solo fumigado a eficiência sobre o crescimento da gravioleira foi de 594%, 847%, 1273% e 1348% em relação aos fungos Entrophosphora colombiana, Gigaspora sp, Gigaspora margarita e Scutellosphora heterogama, respectivamente. Embora o efeito benéfico tenha sido reduzido no solo não fumigado, a eficiência da inoculação ainda foi de 33%, 45%, 51% e 61% nos tratamentos com S. heterogama, Gigaspora sp., E. colombiana e G. margarita, respectivamente.
No segundo corte verificou-se efeito somente das doses de P (p<0,05), para a matéria seca da parte aérea (tabela I), seguindo o modelo quadrático (tabela II). Para maximizar os efeitos da aplicação de P estimou-se a dose 93,5 mg/dm3 de P para um acúmulo máximo de 24,9 g/vaso de MS. No momento do segundo corte o sistema radicular ocupava densamente todo volume do vaso, assim o benefício dos FMAs em aumentar o volume de solo explorado ficaria comprometido, ressaltando a importância da inoculação para o estabelecimento da cultura.
A redução de 700% para 150% em eficiência da inoculação foi observada em mudas de cafeeiro submetidas à inoculação com Gigaspora margarita, na presença de fungos indígenos (Saggin-Júnior e Siqueira, 1996). A eficiência do FMA introduzido no solo natural sem fumigação depende da competitividade sua com o FMA indígeno, no qual está relacionado com a infectividade, a densidade de inóculo e a distribuição dos propágulos de fungos interagentes, a capacidade desses fungos em produzir hifa externa, a velocidade da hifa de colonizar as raízes e a habilidade do fungo introduzido de manter nível de colonização em condição competitiva (Miranda et al., 2005).
Na produção de matéria seca do sistema radicular (MSR), constatou-se efeito de doses de P (p<0,05) e tratamentos micro-biológicos (p<0,05) com independência entre os fatores (tabela I) seguindo modelo quadrático (tabela II). Estimou-se a necessidade da aplicação de 97,0 mg/dm3 de P para alcançar uma produção máxima de 10,7 g/vaso de MSR no tratamento microbiológico controle. De forma independente à dose de P aplicada, a inoculação com Glomus clarum e a com inóculo nativo promoveram incrementos na produção em 2,9 e 1,6 g/ vaso de MSR; e 26,8 e 15,3% a mais, na dose de máxima produção de MSR, respectivamente.
Os resultados de MSR assemelham-se àqueles encontrados por Souza et al. (2000), Santos et al. (2002) e Carneiro et al. (2002). Entretanto, esses autores atribuíram o maior crescimento da parte aérea das leguminosas estudadas, sob inoculação com FMA, ao maior crescimento radicular proporcionado pelos FMAs. No presente estudo, esse fato não se relacionou ao final do período de cultivo, pela não existência de efeito dos tratamentos microbiológicos, para produção de parte aérea, já no segundo corte.
ACÚMULO DE NUTRIENTES
Para acúmulo de nutrientes na matéria seca, Ying Chu et al. (2001) salientaram que uma mesma espécie de fungo micorrízico pode proporcionar respostas diferenciadas em teores de nutrientes nas diferentes plantas hospedeiras e em diferentes condições edafoclimáticas. Santos et al. (2002) enfatizou a maior resposta em acúmulo dos nutrientes de baixa mobilidade no solo como o P, bem como maior acúmulo para o N em leguminosas pelo maior fornecimento de P aos nódulos pela micorriza. Estes autores detectaram ainda, que embora no solo não fumigado a porcentagem de colonização radicular causada por FMAs existentes em plantas sem inoculação não se diferenciasse daquelas causadas por espécies de FMA introduzidos, o aumento em crescimento das mudas de gravioleira com a inoculação de fungos introduzidos ainda foi maior, evidenciando o benefício da inoculação, mesmo no solo não fumigado.
Verificou-se interação significativa entre P x tratamento microbiológico (p<0,05) (tabela I) para o acúmulo de PB no primeiro corte. Estudando o efeito das doses de P para os tratamentos microbiológicos, observa-se que os tratamentos de inoculação sobressaíram-se em relação ao controle, principalmente, nas doses inferiores aplicadas (figura 1b). Os benefícios da inoculação tenderam a diminuir com o aumento das doses, sendo o efeito das doses descrito pelo modelo quadrático em todos os tratamentos microbiológicos (tabela II).
Para o acúmulo de 1497,5 mg/vaso de PB, correspondente à produção máxima obtida no tratamento controle, estimou-se a necessidade da aplicação de 159,6; 12,8 e 53,5 mg/dm3 de P; para os tratamentos controle, Glomus clarum e inóculo nativo, respectivamente. Isso representou uma redução das necessidades em P da ordem de 91,9% quando inoculado com o Glomus clarum, e 66,5% quando inoculado com o inóculo nativo. Os tratamentos de inoculação com FMA proporcionaram efeitos equivalentes à aplicação de 146,8 e 106,1 mg/dm3 de P.
No segundo corte, pela mesma razão apresentada para MS, bem como poderá ser observado para os demais nutrientes, verificou-se efeito somente das doses de P (p<0,05) para o acúmulo de PB (tabela I), seguindo o modelo quadrático (tabela II). Para maximizar os efeitos da aplicação de P, estimou-se a dose 126,6 mg/dm3 para um acúmulo máximo de 3399,0 mg/vaso em PB.
Verificou-se interação significativa entre doses de P x tratamento microbiológico (p<0,05) (tabela I) para o acúmulo de P no primeiro corte. Estudando o efeito das doses de P para os tratamentos microbiológicos, observa-se que os tratamentos de inoculação sobressaíram-se em relação ao controle, principalmente, nas doses inferiores aplicadas (figura 1c). Os benefícios da inoculação tenderam a diminuir com o aumento das doses, sendo o efeito das doses descrito pelo modelo quadrático em todos os tratamentos microbiológicos (tabela II).
Para o acúmulo de 34,6 mg/vaso de P, correspondente ao acúmulo máximo obtido no tratamento controle, estimou-se a necessidade da aplicação de 152,6; 76,6 e 78,3 mg/dm3 de P; para os tratamentos controle, Glomus clarum e inóculo nativo, respectivamente. Isso representou uma redução das necessidades em P, da ordem de 49,8% quando inoculado com o Glomus clarum e 48,6% quando inoculado com o inóculo nativo. Carneiro et al. (2002) encontraram 52 vezes mais fósforo na matéria seca da alfafa quando inoculada com Glomus etunicatum, entretanto estes autores utilizaram solo estéril. Em outro estudo, Carneiro et al. (2008), ao avaliarem a influência do Glomus clarum obtido em bagaço de cana-de-açúcar sobre o estilo-santes em solo natural, concluíram que a inoculação não influenciou a produção de matéria seca, mas foi determinante para o acúmulo superior de P e PB. Percebe-se que apesar da literatura relatar menores valores de produção e acúmulo de nutrientes promovidos pela inoculação com FMA em solo natural, os valores encontrados no presente estudo são significativamente relevantes.
No segundo corte, verificou-se efeito das doses de P (p<0,05) para o acúmulo de P (tabela I), seguindo comportamento quadrático (tabela II). Para maximizar os efeitos da aplicação de P, estimou-se a dose 165,8 mg/dm3 de P para um acúmulo máximo de 81,7 mg/vaso em P.
Assim como os resultados encontrados por Souza et al. (2000) inoculando o Glomus etunicatum associado a doses de P sobre o estilosantes em solo fumigado, as quantidades acumuladas de potássio (K), cálcio (Ca) e enxofre (S), no primeiro corte, não se constatou efeito de interação entre doses de P e tratamentos microbiológicos (p>0,05) (tabela I). Os efeitos foram independentes entre doses de P e tratamentos microbio-lógicos (p<0,05).
Para o potássio (K) estimou-se a necessidade da aplicação de 134,4 mg/dm3 de P para alcançar um acúmulo máximo de 279,8 mg/vaso no tratamento microbiológi-co controle. De forma independente à dose de P aplicada, a inoculação com Glomus clarum e a com inóculo nativo promoveram incrementos em 73,0 e 73,8 mg/vaso; e 26,0 e 26,3% a mais, na dose de máximo acúmulo de K, respectivamente (tabela II).
Para o cálcio (Ca) estimou-se a nece-ssidade da aplicação de 141,9 mg/dm3 de P para alcançar um acúmulo máximo de 153,1 mg/vaso no tratamento microbiológico controle. De forma independente à dose de P aplicada, a inoculação com Glomus clarum e a com inóculo nativo promoveram incrementos em 37,8 e 38,0 mg/vaso; e 24,6 e 24,8% a mais, na dose de máximo acúmulo de Ca, respectivamente (tabela II).
Para o enxofre (S) estimou-se a necessi-dade da aplicação de 132,7 mg/dm3 de P para alcançar um acúmulo máximo de 39,5 mg/ vaso no tratamento microbiológico controle. De forma independente à dose de P aplicada, a inoculação com Glomus clarum e a com inóculo nativo promoveram incrementos em 11,4 e 10,0 mg/vaso; e 29,0 e 25,5% a mais, na dose de máximo acúmulo de S, respectivamente (tabela II).
No segundo corte, o acúmulo de K não foi influenciado pelos tratamentos estudados (p>0,05) (tabela I), sendo de 288,4 mg/vaso o acúmulo geral médio na matéria seca do estilosantes (tabela II). Para o Ca, houve efeito dos tratamentos microbiológicos (p<0,05), independente das doses de P (tabela I). Estimou-se a necessidade da aplicação de 103,6 mg/dm3 de P para atingir um acúmulo máximo de 316,0 mg/vaso no tratamento microbiológico controle. De forma independente à dose de P aplicada, a inoculação com Glomus clarum e a com inóculo nativo promoveram incrementos em 48,3 e 49,7 mg/vaso; e 15,3 e 15,7% a mais, na dose de máximo acúmulo de Ca, respectivamente (tabela II).
Para o enxofre no segundo corte, verificou-se efeito das doses de P (p<0,05) (tabela I) seguindo o modelo quadrático (tabela II). Para maximizar os efeitos da aplicação de P estimou-se a dose 125,9 mg/ dm3 de P para um acúmulo máximo de 112,6 mg/vaso.
Verificou-se interação significativa entre doses de P x tratamento microbiológico (p<0,05) (tabela I) para o acúmulo de Mg no primeiro corte. Estudando o efeito das doses de P para os tratamentos microbiológicos, observa-se que os tratamentos de inoculação sobressaíram-se em relação ao controle, principalmente, nas doses inferiores aplicadas (figura 1d). Os benefícios da inoculação tenderam a diminuir com o aumento das doses, sendo o efeito das doses descrito pelo modelo quadrático em todos os tratamentos microbiológicos (tabela II).
Para o acúmulo de 32,2 mg/vaso de Mg, correspondente à produção máxima obtida no tratamento controle, estimou-se a nece-ssidade da aplicação de 156,3; 50,8 e 68,3 mg/ dm3 de P; para os tratamentos controle, Glomus clarum e inóculo nativo, respectivamente. Isso representou uma redução das necessidades em P da ordem de 67,5% quando inoculado com o Glomus clarum, e 56,3% quando inoculado com o inóculo nativo.
Assim como foi verificado para o Ca, as quantidades acumuladas de Mg, no segundo corte, também tiveram efeito de doses de P e tratamentos microbiológicos de forma independentes (p<0,05) (tabela I). Estimou-se a aplicação de 103,6 mg/dm3 de P para atingir um acúmulo máximo de 125,3 mg/ vaso de Mg no tratamento microbiológico controle. De forma independente à dose de P aplicada, a inoculação com Glomus clarum e a com inóculo nativo promoveram incrementos em 12,0 e 12,7 mg/vaso; e 15,5 e 16,3% a mais, na dose de máximo acúmulo de Mg, respectivamente (tabela II).
Sieverding (1991) relatou que parâmetros fundamentais são necessários de se conhecer para o sucesso no manejo das micorrizas, e que medirão a expectativa de sucesso com a inoculação tanto em aumento de produtividade como eficiência em acúmulo de nutrientes: o grau de heterogeneidade de distribuição dos FMAs nos solos, a variação na concentração de inóculos nativos e da eficiência destes em otimizarem o crescimento das culturas.
COLONIZAÇÃO MICORRÍZICA E DENSIDADE DE ESPOROS
Os tratamentos microbiológicos não influenciaram (p>0,05) a porcentagem de colonização micorrízica do sistema radicular (COL) do estilosantes, sendo apenas influenciada (p<0,05) pelas doses de P (tabela I). Observou-se comportamento linear decrescente, ou seja, um decréscimo médio de 0,1429 unidades percentuais para cada mg/dm3 de P aplicado ao solo (tabela II, figura 1e).
A concentração de propágulos nativos de FMA é correlacionada com a condição de cobertura vegetal no solo, com os tipos de solo (os arenosos em geral têm menores populações de FMA) e com as práticas agronômicas. Menores números de propá-gulos têm sido encontrados em solos com vegetação degradada. Carneiro et al. (1999) verificaram redução no requerimento externo de P pelo estilosantes, capim-braquiária, e, no cultivo consorciado, em função da inoculação micorrízica em um solo degradado por lavagem do horizonte A, com uma estimativa de 9 esporos de FMA nativos por 50 ml de solo.
Provavelmente o aumento na população nativa de FMA, no tratamento controle ao longo do período experimental, tenha contribuído para a igualdade do controle em relação aos tratamentos de inoculação (Glomus clarum e inóculo nativo reinoculado). Como avaliou-se FMA e doses de P em substrato não esterilizado (em condição microbiológica natural), notou-se que, no segundo corte, os efeitos dos tratamentos microbiológicos tenderam à igualdade, com exceção para as quantidades acumuladas de Ca e Mg. Fato este que reforça a suposição de que os resultados, no segundo corte, refletem os resultados de colonização micorrízica e densidade de esporos encontrados ao final do experimento.
Assim como a colonização micorrízica, a densidade de esporos micorrízicos também retrata comportamento semelhante. Verificou-se efeito significativo (p<0,05) apenas para doses de P (tabela I), em comportamento quadrático decrescente (ponto crítico mínimo). Estimou-se a dose de 213 mg/dm3 para uma densidade de esporos mínima de 23,4 esporos (tabela II, figura 1f).
Sieverding (1991) atentou para o fato de que o número de esporos pode ter ou não correlação com o total de propágulos infectivos no solo. Sua taxa pode cair a próximo de zero em culturas a pleno pico de vegetação e com alta infecção de raíz, dependendo do fungo micorrízico, do hospedeiro, das características do solo e das condições climáticas. Este autor relata, ainda, que a taxa de infecção da raiz é uma estimativa da biomassa fúngica interna da raiz. E raramente tem sido encontrada correlação entre a taxa de infecção e propágulos infectivos.
Cavalcante et al. (2002) observaram ausência de interação entre densidade de esporos e espécies de FMA para o crescimento de mudas do maracujazeiro. No entanto, a biomassa seca da parte aérea e área foliar atingiram valores máximos no tratamento com 300 esporos/planta.
Antunes e Cardoso (1990) inocularam 500 esporos/planta de Glomus clarum, e não conseguiram bons resultados com porta-enxertos de citrus. Inoculando apenas 12 esporos/planta de Glomus macrocarpum, Gnekow e Marschner (1989), observaram aumentos significativos na biomassa seca de macieira, seis meses após o plantio. Siqueira et al. (1994) verificaram que a máxima produção da parte aérea do café ocorreu quando foram inoculados 100 esporos/planta e sugeriram que, como em outras culturas, a elevação a partir de certo nível de esporos é desnecessária para obtenção de respostas positivas. Esses resultados revelam que a esporulação possui estreita relação, entre outros fatores, com a espécie hospedeira.
Conclusões
A inoculação com fungos micorrízicos arbusculares para o estabelecimento do estilosantes, em solo não esterilizado, é uma prática promissora utilizando-se principalmente a espécie Glomus clarum.
Bibliografia
Andrade, R.P. e Karia, C.T. 2000. Uso de Stylosanthes em pastagens no Brasil. Em: Evangelista, A.R.; Bernardes, T.F.; Sales, E.C.J. (Eds.). Simpósio de Forragicultura e Pastagens: temas em evidência. NEFOR/UFLA. Lavras. pp. 273-311. [ Links ]
Antunes, V. e Cardoso, E.J.B.N. 1990. O fósforo e a micorriza vesículo-arbuscular no crescimento de porta-enxertos de Citrus cultivados em solo natural. Rev. Bras. Ciên. Solo, 14: 277-282. [ Links ]
Cavalcante, U.M.T., Maia, L.C., Melo, A.M.M. e Santos, V.F. 2002. Influência da densidade de fungos micorrízicos arbusculares na produção de maracujazeiro-amarelo. Pesqui. Agropecu. Bras., 37: 643-649. [ Links ]
Carneiro, R.F.V., Evangelista, A.R., Tonelli, M.T.L. e Reis, S.T. 2002. Inoculação com fungos micorrízicos em alfafa (Medicago sativa L.) em solo com doses crescentes de fósforo. Rev. Ciên. Agrotec., 26: 618-625. [ Links ]
Carneiro, R.F.V., Martins, M.A., Freitas, M.S.M., Detmann, E. e Vasquez, H.M. 2008. Efeito do resíduo agroindustrial e micorríza sobre o crescimento do estilosantes. Arch. Zootec., 57: 259-269. [ Links ]
Carneiro, M.A.C., Siqueira, J.O., Curi, N. e Moreira, M.F.S. 1999. Efeitos da inoculação de fungos micorrízicos arbusculares e da aplicação de fósforo no estabelecimento de forrageiras em solo degradado. Pesqui. Agropecu. Bras., 34: 1669-1677. [ Links ]
Draper, N.R. and Smith, H. 1996. Applied regression analysis. John Wiley de Sous. New York. 407 pp. [ Links ]
Faria, M.P., Siqueira, J.O., Vale, F.R. e Curi, N. 1995. Crescimento de leguminosas arbóreas em resposta a fósforo, nitrogênio, fungo micorrízico e rizóbio.I - Albizia lebbeck (L.) BENTH. Revista Árvore, 19: 293-307. [ Links ]
Fontes, A.G. 2001. Eficiência nutricional e trocas gasosas de espécies florestais em função de doses de fósforo e calagem. Dissertação (Mestrado em Produção Vegetal). Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF). 58 pp. [ Links ]
Gerdemann, J.W. and Nicolson, T.H. 1963. Spores of mycorrhizal endogone species extracted from soil by west sieving and decanting. T. Brit. Mycol. Soc., 46: 235-246. [ Links ]
Giovanetti, M. and Mosse, B. 1980. An evaluation of techniques to measure vesicular-arbuscular mycorrhizal infection roots. New Phytol., 84: 489-500. [ Links ]
Gnekow, M.A. and Marschner, H. 1989. Role of VA-mycorrhiza in growth and mineral nutrition of apple (Malus pumila var. domestica) rootstock cuttings. Plant Soil, 119: 285-293. [ Links ]
Malavolta, E., Vitti, G.C. e Oliveira, S.A. 1989. Avaliação do estado nutricional das plantas: princípios e aplicações. Potafos. Piracicaba. 210 pp. [ Links ]
Moreira, F.M.S. e Siqueira, J.O. 2006. Microbiologia e bioquímica do solo. UFLA. Lavras, MG. 626 pp. [ Links ]
Miranda, J.C.C., Vilela, L. e Miranda, L.N. 2005. Dinâmica e contribuição da micorriza arbuscular em sistemas de produção com rotação de culturas. Pesqui. Agropecu. Bras., 40: 1005-1014. [ Links ]
Oliveira, J.P., Burity, H.A., Lyra, M.C.C.P. e Lira Junior, M.A. 1996. Avaliação da fixação e transferência de nitrogênio na associação gramíneas-leguminosas forrageiras tropicais, através da diluição isotópica do 15N. Rev. Bras. Zootecn., 25: 210-221. [ Links ]
Paulino, V.T., Costa, N.L. e Cardelli, M.A. 1992. Eficiência de fungos micorrízicos vesículo-arbusculares e da adubação fosfatada em Centrosema brasilianum (L.) Benth. Pasturas Tropicales, 14: 14-17. [ Links ]
Rodrigues, L.A., Martins, M.A. e Salomão, M.S.B. 2003. Uso de micorrizas e rizóbio em cultivo consorciado de eucalipto e sesbânia. II Absorção e eficiência de utilização de fósforo e frações fosfatadas. Rev. Bras. Ciên. Solo, 27: 593-599. [ Links ]
Santos, Í.P.A., Pinto, J.C., Siqueira, J.O., Morais, A.R. e Santos, C.L. 2002. Influência do fósforo, micorriza e nitrogênio no conteúdo de minerais de Brachiaria brizantha e Arachis pintoi consorciados. Rev. Bras. Zootecn., 31: 605-616. [ Links ]
Sanggin-Junior, O.J. e Siqueira, J.O. 1996. Micorrizas aebusculares em cafeeiro. Em: Siqueira, J.O. (Ed.). Avanços em fundamentos e aplicação de micorrizas. UFLA/DCS e DCF. Lavras. pp. 203-253. [ Links ]
Souza, R.F., Pinto, J.C., Siqueira, J.O., Curi, N. e Morais, A.R. 2000. Influência de micorriza e fósforo sobre o rendimento de matéra seca e qualidade de A. gayanus e S. guianensis cultivados em um Latossolo. Pasturas Tropicales, 22: 34-41. [ Links ]
Schiavo, J.A. e Martins, M.A. 2002. Produção de mudas de goiabeira (Psidium guajava L.) inoculadas com o fungo micorrízico arbuscular Glomus clarum em substrato agro-industrial. Rev. Bras. Frutic., 24: 519-523. [ Links ]
Schussler, A., Schwarzott, D. and Walker, C. 2001. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol. Res., 105: 1413-1421. [ Links ]
Sieverding, E. 1991. Vesicular-arbuscular mycorrhiza management in tropical agrosystems. Deutsche Gesellschaft fur Techinish Zusamme-narbeit. Eschborn. 371 pp. [ Links ]
Siqueira, J.O., Colozzi-Filho, A. e Saggin-Júnior, O.J. 1994. Efeitos da infecção de plântulas de cafeeiro com quantidades crescentes de esporos do fungo endomicorrízico Gigaspora margarita. Pesqui. Agropecu. Bras., 29: 875-883. [ Links ]
Tebaldi, F.L.H., Silva, J.F.C., Vasquez, H.M. e Thiebaut, J.T.L. 2000. Composição mineral das pastagens das regiões Norte e Noroeste do Estado do Rio de Janeiro. 1. Cálcio, fósforo, magnésio, potássio, sódio e enxofre. Rev. Bras. Zootecn., 29: 603-615. [ Links ]
Vilela, L., Martha Júnior, G.B., Barioni, L.G. e Barcellos, A. 2004. Adubação na recuperação e na intensificação da produção animal em pastagens. Em: Simpósio sobre o manejo da pastagem, 21. Anais... FEALQ. Piracicaba. pp. 425-472. [ Links ]
Ying Chu, E., Moller, M.R.F. e Carvalho, J.G. 2001. Efeitos da inoculação micorrízica em mudas de gravioleira em solo fumigado e não fumigado. Pesqui. Agropecu. Bras., 36: 671-680. [ Links ]
Recibido: 16-10-08.
Aceptado: 16-12-08.

