










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkNutrición Hospitalaria
versão On-line ISSN 1699-5198versão impressa ISSN 0212-1611
Nutr. Hosp. vol.25 no.6 Madrid Nov./Dez. 2010
Relation of C358A polymorphism of the endocannabinoid degrading enzyme fatty acid amide hydrolase (FAAH) with obesity and insulin resistance
Relación del polimorfismo C358A del enzima del sistema endocanabinoide (hidrolasa amida acida) con la obesidad y la resistencia a la insulina
D. A de Luis, M. González Sagrado, R. Aller, O. Izaola, R Conde
Center of Investigation of Endocrinology and Clinical Nutrition, Medicine School and Unit of Investigation. Hospital Rio Hortega. University of Valladolid. Valladolid Spain.
ABSTRACT
Background and aims: Recently, it has been demonstrated that the polymorphism 385 C->A of FAAH (fatty acid amide hydrolase) was associated with overweight and obesity. The aim of our study was to investigate the relationship of missense polymorphism (cDNA 385 C-A) of FAAH gene on obesity anthropometric parameters, cardiovascular risk factors and adipocytokines.
Methods: A population of 279 females with obesity (body mass index >30) was analyzed. An indirect calorimetry, tetrapolar electrical bioimpedance, blood pressure, a serial assessment of nutritional intake with 3 days written food records and biochemical analysis (lipid profile, adipocytokines, insulin, CRP and lipoprotein-a) were performed. The statistical analysis was performed for the combined C385A and A385A as a group and wild type C385C as second group.
Results: One hundred and ninety four patients (69.5%) had the genotype C385C (wild type group) and 76 (27.2%) patients had the genotype C358A or A358A (9 patients, 3.2%) (mutant type group). No differences were detected between groups in anthropometric parameters and dietary intakes. Triglycerides (118.9 ± 59.9 mg/dl vs 107.4 + 51.3 mg/dl;p < 0,05), glucose (100.4 ± 19.9 mg/dl vs 94.8 + 11.5mg/dl; p < 0,05), HOMA (3.74 ± 2.2 vs 3.39 + 2.7; p < 0,05) and interleukine 6 (3.3 ± 1.4 pg/ml vs 1.4 ± 2.1 pg/ml; p < 0,05) were higher in wild type group than mutant type group.
Conclusion: The novel finding of this study is the association of the mutant type group A358C and A358A of FAAH with a better cardiovascular profile (triglyceride, glucose, interleukine 6 and HOMA levels) than wild type group.
Key words: Adipokines. Faah. Insulin resistance. Obesity. Polymorphism. Risk factors.
RESUMEN
Antecedentes y objetivos: Recientemente, se ha demostrado que el polimorfismo 385 C/A, de FAAH (hidrolasa amida de ácidos grasos) se asocia con el sobrepeso y la obesidad. El objetivo de nuestro estudio fue investigar la relación de este polimorfismo del gen de FAAH con parámetros antropométricos, factores de riesgo cardiovascular y adipocitoquinas.
Métodos: Una población de 279 mujeres con obesidad (índice de masa corporal> 30) fue analizada. Se realizaron las siguientes determinaciones; calorimetría indirecta, bioimpedancia eléctrica, presión arterial, una evaluación de la ingesta nutricional de 3 días, así como un análisis bioquímico (perfil lipídico, adipocitoquinas, insulina, proteina C reactiva y lipoproteína-(a)). El análisis estadístico se realizó combinando C385A y A385A como grupo mutante y C385C como grupo salvaje.
Resultados: Un total de 194 pacientes (69,5%) tenían el genotipo C385C (genotipo salvaje) y 76 (27,2%) pacientes tenían el genotipo C358A y 9 pacientes (3,2%) el genotipo A358A, formando estos dos el grupo de genotipo mutante. No se detectaron diferencias entre los grupos en los parámetros antropométricos y la ingesta dietética. Sin embargo los pacientes con genotipo salvaje presentaron valores más elevado de triglicéridos (118,9 ± 59,9 mg/dl vs 107,4 + 51,3 mg/dl; p < 0,05), glucosa (100,4 ± 19,9 mg/dl vs 94,8 + 11,5mg/dl; p < 0,05), HOMA (3,74 ± 2,2 vs 3,39 + 2,7; p < 0,05) y de interleukina-6 (3,3 ± 1,4 pg/ml vs 1,4 ± 2,1 pg/ml; p < 0,05) fueron mayores en el grupo de tipo salvaje que el grupo de tipo mutante.
Conclusión: El principal hallazgo de este trabajo es la asociación del genotipo mutante (A358C y A358A) de FAAH con un mejor perfil cardiovascular (triglicéridos, glucosa, interleucina 6 y HOMA) que los pacientes portadores del genotipo salvaje.
Palabras clave: Adipocitoquinas. FAAH. Resistencia a insulina. Obesidad. Polimorfismo. Factores de riesgo.
Introduction
According to recent surveys, approximately two thirds of the population of Spain is overweight1. The current view of adipose tissue is that of an active secretory organ, sending out and responding to signals that modulate appetite, insulin sensitivity, energy expenditure, inflammation and immunity2.
Herbal Cannabis sativa (marijuana) has been known to have many psychoactive effects in humans including robust increases in appetite and body weight3. Nevertheless, the mechanism underlying cannabinoid neurobiological effects have been recently revealed4. The endogenous cannabinoid system mediates and it is positioned both functionally and anatomically5 to be an important modulator of normal human brain behavior. The main inactivating enzyme of endogenous cannabinoid receptor ligands is fatty acid amide hydrolase (FAAH), which has been identified as the catabolic enzyme capable of inactivating most of the endocannabinoids6.The pharmacological effects of some endocannabinoids appear to be regulated by FAAH activity7, suggesting that FAAH has the role to be a modulating enzyme for human behavior.
A missense polymorphism (cDNA 385 C->A) that predicts a substitution of threonine for a conserved proline residue at aminoacid position 129 (P129T) has been described and is significantly associates with drug abuse8. Recently, Sipe et al9 have been demonstrated that the homozygous FAAH 385 A/A genotype was associated with overweight and obesity. Monteleone et al10 confirmed previously published significant over-representations of the FAAH 385A allele in overweigth/obese subjects. However, in a large study sample (5801 subjects), Jensen et al11 were unable to find association of this polymorphism with overweight or obesity. Considering the evidence that endogenous cannabinoid system plays a role in metabolic aspects of body weight and feeding behavior12, we decided to investigate the association of this FAAH missense polymorphism with obesity and adipocytokines.
The aim of our study was to investigate the relationship of missense polymorphism (cDNA 385 C->A) of FAAH gene on obesity anthropometric parameters, cardiovascular risk factors and adipocytokines.
Subjects and methods
Subjects
A population of 279 females with obesity (body mass index >30) was analyzed in a prospective way. These patients were recruited in a Nutrition Clinic Unit and signed an informed consent. Exclusion criteria included history of cardiovascular disease or stroke during the previous 36 months, total cholesterol > 300 mg/dl, triglycerides > 400 mg/dl, blood pressure > 140/90 mmHg, fasting plasma glucose > 110 mg/dl, as well as the use of sulphonilurea, thiazolidinedions, insulin, glucocorticoids, antineoplasic agents, agiotensin receptor blockers, angiotensin converting enzyme inhibitors and psychoactive medications. Local ethical committee approved the protocol.
Procedure
All patients with a 2 weeks weight-stabilization period before recruitment were enrolled. Weight, blood pressure, basal glucose, c-reactive protein (CRP), insulin, insulin resistance (HOMA), total cholesterol, LDL-cholesterol, HDL-cholesterol, triglycerides blood and adipocytokines (leptin, adiponectin, resistin, TNF alpha, and interleukin 6) levels were measured at basal time. A tetrapolar bioimpedance, an indirect calorimetry and a prospective serial assessment of nutritional intake with 3 days written food records were realized. Genotype of FAAH gene polymorphism was studied.
Genotyping of FAAH gene polymorphism
Oligonucleotide primers and probes were designed with the Beacon Designer 4.0 (Premier Biosoft International®, LA, CA). The polymerase chain reaction (PCR) was carried out with 50 ng of genomic DNA, 0.5 uL of each oligonucleotide primer (primer forward, 5´-CTA TCT GGC TGA CTG TGA GAC TC-3´; primer reverse, 5´-GAG GCA GAG CAT ACC TTG TAG G-3), and 0.25 uL of each probes (wild probe: 5´-Fam-CTG TCT CAG GCC CCA AGG CAG G-BHQ-1-3´) and (mutant probe: 5´-Hex- CTG TCT CAG GCC ACA AGG CAG G-BHQ-1-3´) in a 25 uL final volume (Termociclador iCycler IQ (Bio-Rad®), Hercules, CA). DNA was denaturated at 95oC for 3 min; this was followed by 50 cycles of denaturation at 95oC for 15 s, and annealing at 59.3o for 45 s). The PCR were run in a 25 uL final volume containing 12.5 uL of IQTM Supermix (Bio-Rad®, Hercules, CA) with hot start Taq DNA polymerase. Hardy Weinberg equilibrium was assessed.
Assays
Plasma glucose levels were determined by using an automated glucose oxidase method (Glucose analyser 2, Beckman Instruments, Fullerton, California). Insulin was measured by RIA (RIA Diagnostic Corporation, Los Angeles, CA) with a sensitivity of 0.5mUI/L (normal range 0.5-30 mUI/L)13 and the homeostasis model assessment for insulin sensitivity (HOMA) was calculated using these values14. CRP was measured by immunoturbimetry (Roche Diagnostcis GmbH, Mannheim, Germany), with a normal range of (0-7 mg/dl) and analytical sensivity 0.5 mg/dl. Lipoprotein (a) was determined by immunonephelometry with the aid of a Beckman array analyzer (Beckman Instruments, Calif., USA).
Serum total cholesterol and triglyceride concentrations were determined by enzymatic colorimetric assay (Technicon Instruments, Ltd., New York, N.Y., USA), while HDL cholesterol was determined enzymatically in the supernatant after precipitation of other lipoproteins with dextran sulfate-magnesium. LDL cholesterol was calculated using Friedewald formula.
Adipocytokines
Resistin was measured by ELISA (Biovendor Laboratory, Inc., Brno, Czech Republic) with a sensitivity of 0.2 ng/ml with a normal range of 4-12 ng/ml15. Leptin was measured by ELISA (Diagnostic Systems Laboratories, Inc., Texas, USA) with a sensitivity of 0.05 ng/ml and a normal range of 10-100 ng/ml16. Adiponectin was measured by ELISA (R&D systems, Inc., Mineapolis, USA) with a sensitivity of 0.246 ng/ml and a normal range of 8.65-21.43 ng/ml17. Interleukin 6 and TNF alpha were measured by ELISA (R&D systems, Inc., Mineapolis, USA) with a sensitivity of 0.7 pg/ml and 0.5 pg/ml, respectively. Normal values of IL6 was (1.12-12.5 pg/ml) and TNFalpha (0.5-15.6 pg/ml)17-19.
Indirect calorimetry
For the measurement of resting energy expenditure, subjects were admitted to a metabolic ward. After a 12 h overnight fast, resting metabolic rate was measured in the sitting awake subject in a temperature-controlled room over one 20 min period with an open-circuit indirect calorimetry system (standardized for temperature, pressure and moisture) fitted with a face mask (MedGem;Health Tech, Golden, USA), coefficient of variation 5%. Resting metabolic rate (kcal/day) and oxygen consumption (ml/min) were calculated20.
Anthropometric measurements
Body weight was measured to an accuracy of 0.5 Kg and body mass index computed as body weight/(height2). Waist (narrowest diameter between xiphoid process and iliac crest) and hip (widest diameter over greater trochanters) circumferences to derive waist-to hip ratio (WHR) were measured, too. Tetrapolar body electrical bioimpedance was used to determine body composition with an accuracy of 5 g21. An electric current of 0.8 mA and 50 kHz was produced by a calibrated signal generator (Biodynamics Model 310e, Seattle, WA, USA) and applied to the skin using adhesive electrodes placed on right-side limbs. Resistance and reactance were used to calculate total body water, fat and fat-free mass.
Blood pressure was measured twice after a 10 minutes rest with a random zero mercury sphygmomanometer, and averaged.
Dietary intake and habits
Patients received prospective serial assessment of nutritional intake with 3 days written food records. All enrolled subjects received instruction to record their daily dietary intake for three days including a weekend day. Handling of the dietary data was by means of a personal computer equipped with personal software, incorporating use of food scales and models to enhance portion size accuracy. Records were reviewed by a registered dietitian and analyzed with a computer-based data evaluation system. National composition food tables were used as reference22.
Statistical analysis
Sample size was calculated to detect differences over 2 kg in body weight with 90% power and 5% significance (n=80, in each group). The results were expressed as average ± standard deviation. The distribution of variables was analyzed with Kolmogorov-Smirnov test. Quantitative variables with normal distribution were analyzed with a two-tailed Student´s-t test. Non-parametric variables were analyzed with the U-Mann-Whitney test. The statistical analysis was performed for the combined C385A and A385A as a group and wild type C385C as second group. A p-value under 0.05 was considered statistically significant.
Results
Two hundred and seventy nine patients gave informed consent and were enrolled in the study. The mean age was 44.8+12 years and the mean BMI 35.4 ± 6.1. All subjects were weight stable during the 2 weeks period preceding the study (body weight change, 0.28 ± 0.1 kg).
One hundred and ninety four patients (69.5%) had the genotype C358C (wild type group) and 76 (27.2%) patients had the genotype C358A or A358A (9 patients,3.2%) (mutant type group). Age was similar in both groups (wild type: 44.70 ± 15.5 years vs mutant group: 45.5 ± 15.2 years:ns).
Table I shows the anthropometric variables. No differences were detected between groups.
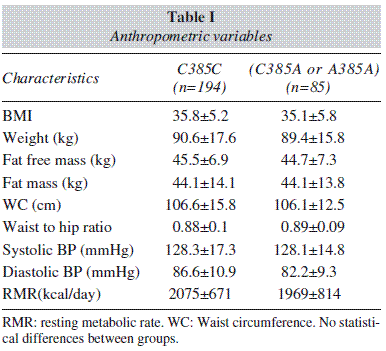
Table II shows the classic cardiovascular risk factors. Triglycerides, glucose and HOMA values were higher in wild type group than mutant type group.
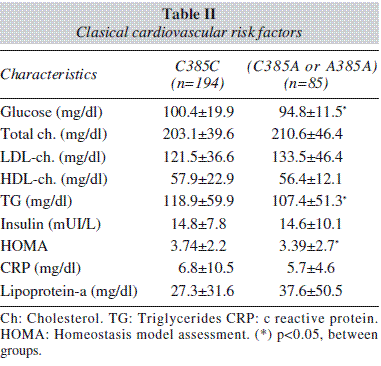
Table III shows nutritional intake with 3 days written food records. No statistical differences were detected in caloric, carbohydrate, fat, and protein intakes. Aerobic exercise per week was similar in both groups.
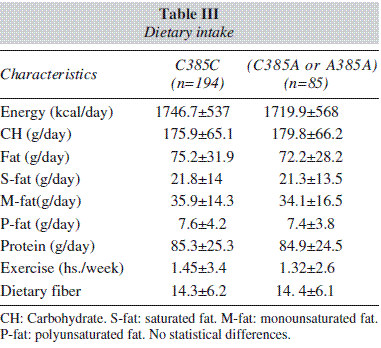
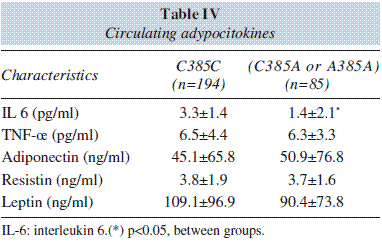
Table IV shows levels of adipocytokines. IL-6 levels were higher in wild type group than mutant type group. No differences were detected between both groups in other serum adipocytokine levels.
Discussion
Since genetic influences are thought to play a greater role in obesity, identification of obesity genes may be more easily detected in the former. A common C358A single nucleotide polymorphism (SNP) of the FAAH results in a missense mutation producing a FAAH with defective expression23. Studies of the C358A variant and obesity from various authors have yeldied conflicting results8-11.
In our study, the percentage of AA genotype (3.2%) was similar than others, for example; 3.7%9, 2.3%25 and 1%10. The percentage of AC genotype (27.2%) was similar than other studies; 28.1%9, 24.1%25 and 36.5%10, too.
In our design, we investigated the effect of FAAH genetic variation on weight, and also on metabolic parameters. The novel finding of this study is the association of the C385C FAAH genotypes with higher levels of IL6, glucose, HOMA and triglycerides than C385A and A385A FAAH genotypes than wild type group, without differences in anthropometric parameters related with obesity.
An association between this polymorphism and metabolic profile has been described. Interestingly, Aberle et al24 have shown that carriers of the A allele had a significantly greater decrease in the total cholesterol and triglycerides as compared to wild type when folloging a low fat diet.
The lack of association between this polymorphism and anthropometric parameters has been described by other authors. The results of our study agree with those of Jensen et al11 or Papazoglou et al25 and contrast with those of Sipe et al9. It is noteworthy that when BMI was evaluated as a continous variable in subjects, the median BMI was significantly higher in subjects with the A385A genotype compared to the median BMI of the other subjects9. The effects of the FAAH A385A could result in the upregulation of natural reward behaviours such as consumption of sweets and palatable food associated with overweigth and obesity. Under normal circumstances, brain endocanabinoid signalling may exert tonic control on the levels of certain neurotransmitters. The mesolimbic addition and reward/craving circuit including the medial forebrain bundle projections to the nucleus accumbens shows a high correlation of FAAH enzyme expression and CB1 receptor density26. However, the inconsistencies between association studies may reflect the complex interactions between multiple population-specific genetic and environmental factors. Perhaps, these different results could be explained by inclusion criteria of subjects in previous studies of the literature. These previous studies would require composition analysis of the diet to determine whether dietary components could be responsible for the lipid profile modifications. In our study dietary intake did not show statistical differences between groups, in this way our datas have been controlled by dietary intake and previous discrepancies could be explain by this uncontrolled factor (dietary intake).
Accumulating evidence indicates that the endogenous cannabinoid system is an essential homeostatic regulator of energy balance and weight via central appetite-stimulating mechanisms as well as peripheral lipogenesis27. The anatomical convergence of CB1 receptors and FAAH enzyme activity capable of regulating endocannabinoid tone supports the notion that FAAH is positioned for regulation of endocannabinoid levels that could influence craving and reward behaviors through the relevant neuronal circuitry. Patients with this polymorphism have half the FAAH enzymatic activity and protein expression when compared with wild-type subjects23. This provide a link between the consequences of this polymorphism and the present epidemiological study indicating that the FAAH A385A polymorphism may play a role in decrease or increase metabolic risk associated to obesity.
In our study, the elevation of inflammation markers (IL-6) in patients with wild type genotype could be related with an altered postprandial response of fatty acids absorption (no measured in our design). Elevated free fatty acids increases the acummulation of triglycerides in the adipocyte, related with imbalance of lipoprotein lipase activity and overproduction of adipokines such as IL-6. The lack of association of this proimflammatory state with fat mass or BMI could indicate the existence of complex unmeasured gene-gene or gene-environment interactions that may enhance metabolic abnormalities in obese patients.
In conclusion, the novel finding of this study is the association of the mutant type group A385C and A385A of FAAH with a better cardiovascular profile (triglyceride, glucose, interleukine 6 and HOMA levels) than wild type group. Further studies are needed to elucidate this complex relationship in an independent way of body mass index.
References
1. Aranceta J, Perez Rodrigo C, Serra Majem L. Prevalencia de la obesidad en España: estudio SEEDO 97. Med clin (Barc) 1998; 111: 441-445. [ Links ]
2. Matsuzawa Y. Adipocytokines: Emerging therapeutic targets. Current Atherosclerosis Reports 2005; 7: 58-62. [ Links ]
3. Ameri A. The effects of cannabinnoids on the brain. Prog Neurobiol 1999;58:315-348. [ Links ]
4. Brown SM, Warger- Miller J, Mackie K. Cloning and molecular characterization of the rat CB2 cannabinoid receptor. Biochim Biophys Acta 2002; 1576: 255-264. [ Links ]
5. Fride E. Endocannabinoids in the central nervous system an overview. Prostaglandins Leukot Essent Fatty acids 2002; 66: 221-233. [ Links ]
6. Deustch DG, Ueda N, Yammamoto S. The fatty acid amide hydrolase (FAAH) 1. Prostaglandins Leukot Essent Fatty Acids 2002; 66: 201-210. [ Links ]
7. Lichtman AH, Hawkins EG, Griffin G, Cravatt BF. Pharmacological activity of fatty acid amides is regulated, but not mediated, by fattu acid amide hydrolase in vivo. Pharmacol Exp Ther 2002; 302: 73-79. [ Links ]
8. Sipe JC, Chiang K, Gerber AL, Beutler E, Cravatt BF. A Missense mutation in human fatty acid amide hydroxylase associated with problem drug abuse. Proc Natl Acad Sci USA 2002; 99: 8394-8399. [ Links ]
9. Sipe JC, Waalen J, Gerber A, Beutler E. Overweight and obesity associated with a missense polymorphism ain fatty acd amide hydrolase (FAAH). Inte J of Obes 2005; 29: 755-759. [ Links ]
10. Monteleone P, Tortorella A, Martiadis V, Filippo C, Canestrelli B, Maj M. The CDNA 385 C to A missense polymorphism of the endocannabinoid degrading enzyme fatty acid amide hydrolase (FAAH) os associated with overweight/obesity but not with binge eating disorder in overweight/obese women. Psychoneuroendocrinology 2008; 33: 546-550. [ Links ]
11. Jensen D, Andreasen C, Andersen M, Hansen L, Eiberg H, Borch-Johnsen K, Jorgensen T, Hansen T, Pedersen O. The functional pro129Thr variant of the FAAH gene is not associated with various fat accumulation phenotypes ina a populationbased cohort of 5801 whites. J Mol med 2007; 85: 445-449. [ Links ]
12. Harrold JA, Williams G. The cannabinoid system; a role in both the homeostatic and hedonic control of eating? Br J Nutr 2003; 90: 729-734. [ Links ]
13. Duart MJ, Arroyo CO, Moreno JL. Validation of a insulin model for the reactions in RIA. Clin Chem Lab Med 2002; 40: 1161-1167. [ Links ]
14. Mathews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher Df. Homesotasis model assessment: insulin resistance and beta cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985; 28: 412-414. [ Links ]
15. Pfutzner A, Langefeld M, Kunt T, Lobig M. Evaluation of human resistin assays with serum from patients with type 2 diabetes and different degrees of insulin resistance. Clin Lab 2003; 49: 571-576. [ Links ]
16. Meier U, Gressner M. Endocrine regulation of energy metabolis: review of pathobiochemical and clinical chemical aspects of leptin, Ghrelin, adiponectin, and resistin. Clinical Chemstry 2004; 50: 1511-1525. [ Links ]
17. Suominen P. Evaluation of an enzyme immunometric assay to measure serum adiponectin concentrations. Clin Chem 2004; 50: 219-221. [ Links ]
18. Lubrano V, Cocci F, Battaglia D, Papa A. Usefulness of high-sensitivity IL6 measurmentfor clinical characterization of patients with coronary artery disease. J Clin Lab Anal 2005; 19: 110-114. [ Links ]
19. Khan SS, Smith MS, Reda D, Suffredini AF, Mc Coy JP. Multiplex bead array assays for detection of soluble cytokines: comparisons of sensitivity and quantitative values among kits from multiple manufactures. Cytometry B Clin Cytom 2004; 61: 35-39. [ Links ]
20. Feurer ID, Mullen JL. Bedside measurement of resting energy expenditure and respiratory quotient via indirect calorimetry. Nutr Clin Pract 1986; 1: 43-49. [ Links ]
21. Pichard C, Slosman D, Hirschel B, Kyle U. Bioimpedance analysis: an improved method for nutritional follow up. Clin Res 1993; 41: 53. [ Links ]
22. Mataix J, Mañas M. Tablas de composición de alimentos españoles. Ed: University of Granada, 2003. [ Links ]
23. Chiang KP, Gerber AL, Sipe JC, Cravatt BF. Reduced cellular expresion and activity of the P129T mutant fatty acid amide hydrolase: evidence for a link between defects in the endocannabinoid system and problem drug use. Hum Mol Genet 2004; 13: 2113-2119. [ Links ]
24. Aberle J, Fedderwitz I, Klages N, George E, Beil FU. Genetic variation in two proteins of the endocannabinoid system and their influence on body mass index and metabolism under low fat diet. Horm Metab Res 2007; 39: 395-397. [ Links ]
25. Papazoglou D, Panagopoulos I, Papanas N, Gioka T, Papadopoulos T, Papathanasiou P, Kaitozis O et al. The FAAH Pro129Thr polymorphism is not asociated with severe obesity in Greek Subjects. Horm Metab Res 2008; 40: 907-910. [ Links ]
26. Egertova M, Cravatt BF, Elphick MR. Comparative analysis of FAAH and CB1 canabinoid receptor expression in the mouse brain:evidence of a widespread role for fatty acid amide hydrolase in regulation of endocannabinoid signalling. Neuroscience 2003; 119: 481-496. [ Links ]
27. Cota D, Marsicano G, Lutz B, Vicennatu V, Stalla GK, Pasqualli R, Paggotto U. endogenous cannabinoid system as a modulator of food intake. Int J Obes Relat Metab Disor 2003; 27: 289-301. [ Links ]
![]() Correspondence:
Correspondence:
D. A de Luis
Professor Associated of Nutrition
Executive Director of Institute of Endocrinology and Nutrition.
Medicine Schooll. Valladolid University.
C/ Los perales 16
47130 Simancas (Valladolid), Spain.
E-mail: dadluis@yahoo.es
Recibido: 8-VI-2010.
1a Revisión: 12_VIII-2010.
Aceptado: 11-IX-2010.

