










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkNutrición Hospitalaria
versión On-line ISSN 1699-5198versión impresa ISSN 0212-1611
Nutr. Hosp. vol.28 no.2 Madrid mar./abr. 2013
https://dx.doi.org/10.3305/nh.2013.28.2.6310
ORIGINAL
Influence of early environmental factors on lymphocyte subsets and gut microbiota in infants at risk of celiac disease; the PROFICEL study
Influencia de factores ambientales tempranos sobre las subpoblaciones de linfocitos y la microbiota intestinal de niños con riesgo de desarrollar enfermedad celíaca; el Estudio PROFICEL
Tamara Pozo-Rubio1, Giada de Palma2, Jorge R. Mujico1, Marta Olivares2, Ascensión Marcos1, María Dolores Acuña3, Isabel Polanco4, Yolanda Sanz2 and Esther Nova1
1Immunonutrition Group. Department of Metabolism and Nutrition. Institute of Food Science. Technology and Nutrition (ICTAN). Spanish National Research Council (CSIC). Madrid. Spain.
2Microbial Ecophysiology and Nutrition Group. Institute of Agrochemistry and Food Technology (IATA). Spanish National Research Council (CSIC). Valencia. Spain.
3Gastroenterology Unit. Hospital Infantil Universitario Niño Jesús. Madrid. Spain.
4Gastroenterology and Paediatric Nutrition Service. Hospital Universitario La Paz. Madrid. Spain.
Supported by grants AGL2007-66126-C03-01/ALI, AGL2007-66126-C03-02/ ALI and AGL2007-66126-C03-03/ALI, and Consolider Fun-C-Food CSD2007-00063, from the Spanish Ministry of Science and Innovation and grants 200570F0091 and 200570F0093 from CSIC.
ABSTRACT
Introduction: It is known that the HLA genotype can explain about a 40% of the genetic risk of celiac disease (CD), thus, other genetic predisposing factors as well as factors that subtly modulate T cell activation and differentiation need to be studied. This includes environmental factors that are currently believed to impact on the immune system and gut microbiota development.
Aim: To assess the associations between early environmental factors (EEF), lymphocyte subsets, and intestinal microbiota composition in infants at familial risk for CD.
Study design: Prospective observational study.
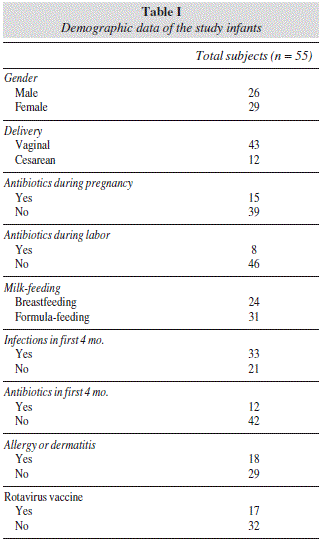
Subjects: Fifty-five 4 month-old infants with at least a first-degree relative suffering CD. Infants were classified according to type of delivery, mother's antibiotic intake during pregnancy and during labor, milk-feeding practices, early infections and antibiotic intake, rotavirus vaccine administration, and allergy incidence within the first 18 months of life.
Methods: Lymphocyte subsets and gut microbiota composition were studied at the age of 4 months.
Results: Formula feeding and infant's infections were associated with higher CD3+, CD4+, CD4+CD38+, CD4+CD28+ and CD3+CD4+CD45RO+ counts (P0.01). Infant s infections were also associated with higher CD4+CD25+, CD4+HLA-DR+ and NK cell counts (P0.01). Cesarean delivery and rotavirus vaccine administration were associated with lower percentage of CD4+CD25+ cells. Infant's antibiotic intake was associated and correlated with lower counts of Bifidobacterium longum and higher counts of Bacteroides fragilis group.
Conclusions: Infant s infections and antibiotic intake in the first 4 months of life are the EEF more strongly and/or frequently associated to lymphocyte subpopulations and microbiota composition, respectively, in infants at risk of CD.
Key words: Lymphocyte subpopulations. Gut microbiota. Infants. Early environmental factors. Celiac disease's risk.
RESUMEN
Introducción: Es bien sabido que el genotipo HLA puede explicar un 40% del riesgo genético de enfermedad celíaca, por lo que otros factores de predisposición genéticos así como factores que sutilmente modulen la activación y diferenciación de células T necesitan ser estudiados. Esto incluye factores ambientales, de los que se cree actualmente que ejercen un efecto sobre el desarrollo del sistema inmune y la microbiota intestinal.
Objetivo: Evaluar las asociaciones entre factores ambientales tempranos, las subpoblaciones de linfocitos y la composición de la microbiota intestinal en niños con riesgo familiar de enfermedad celíaca.
Diseño del estudio: Estudio prospectivo observacional
Sujetos: 55 niños de 4 meses de edad con al menos un familiar celíaco de primer grado. Los niños fueron clasificados de acuerdo al tipo de parto, ingesta materna de antibióticos durante el embarazo y durante el parto, tipo de lactancia, infecciones tempranas y toma de antibióticos, administración de la vacuna de rotavirus, y incidencia de alergias dentro de los 18 primeros meses de vida.
Métodos: Las subpoblaciones de linfocitos y la composición de la microbiota intestinal fueron estudiadas a la edad de 4 meses.
Resultados: La lactancia de fórmula y las infecciones tempranas se asociaron con un mayor número absoluto de células CD3+, CD4+, CD4+CD38+, CD4+CD28+ y CD3+CD4+ CD45RO+ (P0.01). El parto por cesárea y la administración de la vacuna de rotavirus se asociaron a un menor porcentaje de células CD4+CD25+. La toma temprana de antibióticos se asoció y correlacionó con menor número de Bifidobacterium longum y mayor número de Bacteroides fragilis.
Conclusiones: Las infecciones y la toma de antibióticos en los primeros 4 meses de edad son los factores ambientales tempranos más fuertemente y/o frecuentemente asociados a las subpoblaciones de linfocitos y la composición de la microbiota, respectivamente, en niños con riesgo de enfermedad celíaca.
Palabras clave: Subpoblaciones de linfocitos. Microbiota intestinal. Infantes. Factores ambientales tempranos. Riesgo de enfermedad celíaca.
Abbreviations
CD: Celiac disease.
EEF: Early environmental factors.
BF: Breast-fed (infants).
FF: Formula-fed (infants).
PBMCs: Peripheral blood mononuclear cells.
OVA: Ovoalbumin.
Introduction
Celiac disease (CD) is an immune-mediated enteropathy triggered by the ingestion of gluten in genetically susceptible individuals. It is generally accepted that CD is a T-cell mediated disease, in which gliadin derived peptides activate lamina propria infiltrating T lymphocytes. The subsequent release of pro-inflammatory cytokines, in particular y-interferon, leads to a profound tissue remodeling. However, an innate immune response with the participation of IL-15 also seems to be involved in the disease development.1,2 This is a complex disorder, with environmental and genetic factors contributing to its etiology. The major genetic risk factor in CD is represented by HLA-DQ genes which account for approximately 40% of the genetic risk for CD. Approximately 90-95% of CD patients express HLA-DQ2 heterodimers, and the remaining 5-10% of patients express the HLA-DQ8 heterodimer.2 However, only a subset of individuals that express HLA-DQ2 and HLA-DQ8 heterodimers develop CD and, among them, some individuals develop CD very early in infancy after their first exposure to gluten, and others in adulthood; the reasons for these variations are still unclear.2,3 Thus, HLA-DQ2/8 heterodimers are necessary but not sufficient, and other genetic and environmental factors must be involved in CD development.3 Gluten is the main environmental factor responsible for the signs and symptoms of the disease, but also, other environmental elements might play a role in the disease risk.4
The effect of environmental factors on disease risk is relevant at the early stages of life when the immature neonate's gut undergoes the process of microbiota exposure and establishment, and the immune system acquires full competence and tolerance to non-harmful antigens 5. A particular feature of the neonatal immune system is the presence of a wide pool of naive T cells waiting to participate in primary immune responses. In addition, intestinal antigen exposure during neonatal life, and the establishment of the microbiota, influences appropriate adult immune responses.6 If appropriate immune tolerance is not established in early life and maintained throughout life, this represents a risk factor for the development of inflammatory, autoimmune, and allergic diseases.7 CD development is also likely to be influenced by factors that modulate the development and maturation of the immune system in infants.
There are several factors influencing the colonization pattern and establishment of microbiota in infants. The microbes colonizing the infant's gut come primarily from the mother's vaginal and perineal microbiota if vaginally delivered, and from the feeding route, breast feeding or bottle feeding. Thus, the colonization pattern is influenced by delivery and feeding modes, and the exposure to pathogens and antimicrobials.8
As early events during extrauterine life allow the maturation of the immune system, the peripheral lymphocyte subsets exhibit certain changes. Blood cell analysis shows marked lymphocytosis at birth and at later stages of life compared to adulthood. An increase in T and B lymphocytes occurs during the first weeks of life while NK cells decline sharply directly after birth.9,10 A high CD4/CD8 ratio in babies up to two years of age has been found compared to the values observed in normal adults, and this is mainly due to the large and expanding pool of naïve T CD4+ cells present during the first year of life.11,12 The proportion of activated T cells like CD3+CD25+ and CD3+HLA-DR+ are lower in newborns than in adults.13 In contrast, the proportions of CD8+CD28+, CD8+CD38+, and CD4+CD38+ cells are higher in neonates.13,14 There are only a limited number of studies that have addressed the influence of milk-feeding practices15-20 or type of labor or delivery21-23 on peripheral lymphocyte subsets or the immunocompetence of lactating children.
Given the important role that the maturation of the immune system can play as a predisposing or protective factor to disease development later in life, we planned to assess the effect of type of delivery, gender, mother's antibiotic intake during pregnancy, mother's antibiotic administration during labor, milk-feeding practices, infections and antibiotic intake in the first 4 months of life, and rotavirus vaccine administration on the levels of lymphocyte subsets and their activation markers, and on the gut microbiota composition in infants at risk of developing CD.
Materials and methods
Subjects and information on environmental factors
This study was conducted in 4 month-old infants who are first-degree relatives of CD patients (at least one parent or sibling affected with CD). A total of 55 infants with these characteristics, born between October 2006 and the end of 2010 were recruited in the University Hospital La Paz and University Hospital Niño Jesús of Madrid. Questions about mode of delivery, infections, vaccinations, feeding practices, clinical symptoms, and medical treatments were answered by the parents at the infant's age of 1 and 4 months in clinical visits with the paediatric gastroenterologist. To facilitate the collection of this information, a diary was given to the mothers upon recruitment (before delivery). On the basis of these questionnaires, the 55 infants were divided according to the following variables that were selected as potential influential factors of immune system development and gut microbiota composition: type of delivery (vaginal/cesarean), mother's antibiotic intake during pregnancy (yes/no), mother's antibiotic administration during labor (yes/no), milk-feeding practices (breast-fed (BF) infants, including those which had been exclusively breast-fed until the age of 4 months, and formula-fed (FF) infants, including babies receiving milk formula, either alone or in alternation with breast milk at the moment of analysis), infections in the first 4 months of life (yes/no), antibiotic intake in the first 4 months of life (yes/no), rotavirus vaccine administration (yes/no), and allergies or dermatitis suffering within the first 18 months of life (yes/no) (table I). The study was approved by the Ethic Committee of the University Hospital La Paz and the University Hospital Niño Jesús of Madrid, and written informed consent was obtained from the parents of infants included in the study. This group of infants was a subset of those recruited in 8 Spanish hospitals for the PROFICEL study.20
Blood analyses
At the age of 4 months, peripheral blood samples were drawn from the infants and collected in BD VacutainerTM tubes containing K3EDTA. Immediately after, 1 mL of blood was mixed with an equal volume (1 mL) of preservative solution (Streck Cell PreservativeTM CE. Streck, USA) and sent to our laboratory at 4-8o C within 2-6 days for a centralised processing and flow cytometry analysis. Blood extractions were always performed previous to the standardized 4 month vaccination in the Spanish vaccination schedule. Complete blood counts and differential counts were performed in situ at the corresponding enrolling centers by automated instrumentation. Blood samples were not taken if the infant presented an ongoing infection or had received a vaccination in the prior six weeks.
Flow cytometry analysis
Aliquots of blood mixed with the preservative-solution (150 L) were incubated for 30 minutes at room temperature and in the dark with 20 L of fluorochrome conjugated-monoclonal antibodies specific for CD3+ (CD3-APC), CD4+ (CD4-PerCP-Cy5.5), CD8+ (CD8-PerCP-Cy5.5), CD45RA+ (CD45RA-FITC), CD45RO+ (CD45RO-PE), CD25+ (CD25-FITC), HLA-DR+ (HLA-DR-FITC), CD38+ (CD38-PE), in quadruple immunostainings and 100 pL aliquots were incubated with the multitests CD3-FITC/CD16+56-PE/CD45-PerCP-Cy5.5/CD19-APC and CD3-FITC/CD8-PE/CD45-PerCP-Cy5.5/CD4-APC. All the monoclonal antibodies were purchased from Becton Dickinson (Sunnyvale, CA, USA). After incubation, samples were lysed with the BD FACSTM Lysing Solution (Becton Dickinson) following the manufacturer's protocol. The samples were analysed with FACSCal-ibur Flow Cytometer (four-color, dual-laser, Becton Dickinson). The lymphogate was defined on the forward and side scatter patterns of lymphocytes. The analysis protocol gated on lymphocytes stained with PerCP and/or APC and the selected population was then analysed with the 2 remaining colours (FITC and PE) to obtain cell percentages expressing the specific antigens. Cell subset counts were obtained by multiplying subset percentages by anchor marker counts, the later resulting by multiplying subset percentage by the absolute lymphocyte count.
Faecal sampling and DNA extractions
Stool samples were collected from 44 subjects at 4 months of age and frozen at -20oC immediately. Samples (1 g) were diluted 1:10 (w/v) in PBS (pH 7.2) and homogenized by thorough agitation in a vortex. Aliquots were used for DNA extraction using the QIAamp DNA stool Mini kit (Qiagen, Hilden, Germany) following the manufacturer's instructions. DNA extractions from different pure cultures of reference strains were done following the same protocol.
Quantitative PCR (qPCR) analysis of faecal bacteria
The following bacterial groups were analysed: Bifidobacterium spp., Lactobacillus group, Staphylococcus spp., Bacteroides fragilis group, Clostridium coccoides-Eubacterium rectali group, Clostridium leptum group, Escherichia coli, and the following Bifidobacterium spp.: B. longum, B. breve, B. bifidum, B. adolescentis, B. catenulatum, B. angulatum, B. infantis, B. lactis, and B. dentium. qPCR was used to quantify the different bacterial groups in faeces using genus-, group- and species-specific primers as previously described.24,25 Briefly, PCR amplification and detection were performed with an ABI PRISM 7000-PCR sequence detection system (Applied Biosystems, UK) using SYBR® Green PCR Master Mix (Super-Array Bioscience Corporation, USA). The bacterial concentration from each sample was calculated by comparing the Ct values obtained from standard curves of reference strains. Standard curves were created using serial 10-fold dilutions of pure culture DNA corresponding to 102 to 109 cells, as determined by microscopy counts after staining with 4',6-diamino-2-phenylindole in an epifluorescence microscope (Olympus BX51, Tokio, Japan).
Statistics
Associations of perinatal and early environmental factors (EEF) with lymphocyte subset values and bacteria counts were analysed using multiple linear regression models. In addition, logistic regression analyses were used to examine the effect of the environmental factors on the prevalence of colonization (colonized opposed to non-colonized). Previously, data with a non-normal distribution were transformed logarithmically to a normal distribution. Type of delivery, mother's antibiotic intake during pregnancy, mother's antibiotic administration during labor, milkfeeding practices, infections and antibiotic intake in the first 4 months of life, rotavirus vaccine administration, and allergy incidence were included in the models. To limit the chance of falsely rejecting the null hypothesis (no association) as a result of multiple testing, a P-value lower than 0.010 was considered statistically significant in multiple linear regression analysis. Spearman's test was performed to assess possible correlations between exposures to the different individual perinatal and environmental factors and lymphocyte subsets, and also between the same factors and microbiota composition. Finally, correlations between lymphocyte subsets and faecal bacteria counts were equally performed. A P-value less than 0.050 was considered statistically significant.
Results
Lymphocytes subsets
The associations of perinatal and EEF with the percentage and counts of the lymphocyte subsets under study, as determined in the linear regression analysis adjusted for gender, are presented in table II.
Formula-feeding was associated with higher absolute counts of total lymphocytes, T cells (CD3+), helper T cell (CD4+), CD4+CD38+, and CD4+CD28+ cells. T test analyses confirmed these results (P values < 0.05 for these four subsets), and also showed that the percentage of CD4+CD25+ cells was higher in BF infants (11.1 ± 2.9 vs. 9.3 ± 3.3; P < 0.05) while CD4+CD38+ cell percentage was lower compared to FF infants (91.1 ± 19.5 vs. 96.6 ± 1.0; P ≤ 0.05).
Infant's infections in the first 4 months of age were also associated with higher absolute counts of CD3+, CD4+, CD4+CD38+, and CD4+CD28+ cells (table III).
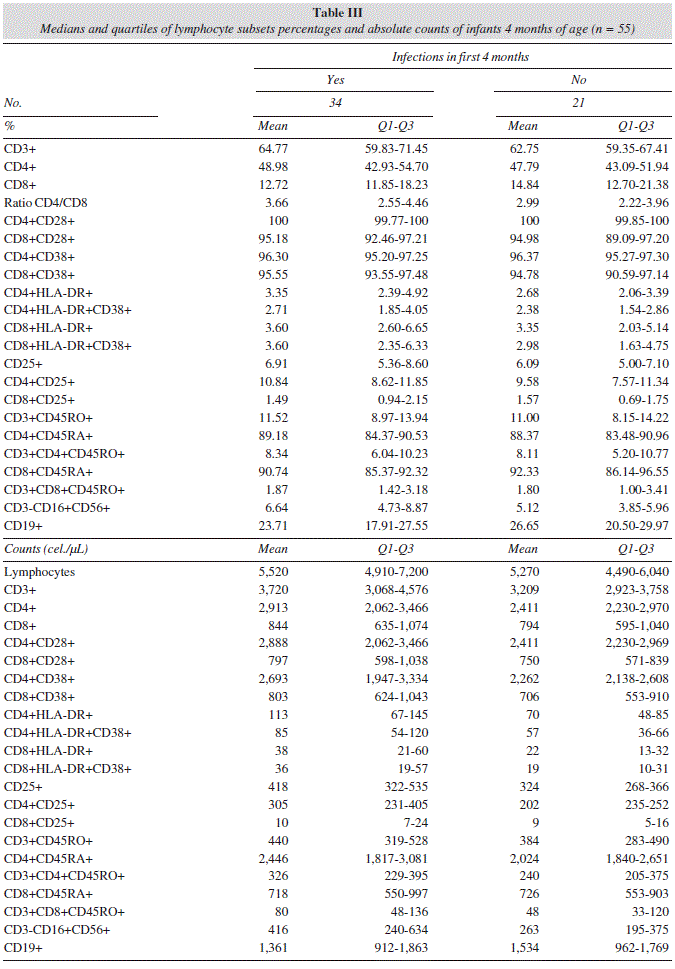
Regarding activation markers which are not constitutively expressed, cesarean delivery and rotavirus vaccine administration were associated with a lower percentage of activated T helper lymphocytes that express CD25+ (CD4+CD25+), in contrast, infants whose mothers used antibiotics during pregnancy were associated with a higher percentage of these cells. Infant's infections in the first 4 months of life were associated with higher absolute counts of CD4+CD25+ (table III).
Formula-feeding and infant's infections were associated with higher absolute counts of memory T helper lymphocytes (CD3+CD4+CD45RO+), and infant's antibiotic intake in the first 4 months was associated with lower absolute counts of these cells.
Infant's infections were associated with higher absolute counts of activated T helper lymphocytes HLA-DR+ (CD4+HLA-DR+) (table III).
Finally, infant's infections and antibiotic administration in mothers during labor were associated with higher percentage and absolute counts of NK cells (CD3-CD16+CD56+), and future allergy incidence was associated with a lower percentage of NK cells.
Spearman's test carried out to analyse the correlations between lymphocyte subsets and bacterial genera, groups and species, did not show significant results.
Gut microbiota composition
The composition of the fecal microbiota of 44 infants was analysed at 4 months of age (table IV). The remaining 11 infants did not collect their stool samples.
The results of the linear regression analysis of bacterial counts with respect to environmental factors are shown in table IV. Infants born by cesarean delivery showed associations with lower counts of B. catenulatum, and higher counts of B. angulatum (table V). Spearman correlations showed that infants born by cesarean delivery were correlated with lower counts of B. catenulatum (R = -0.514; P = 0.010).
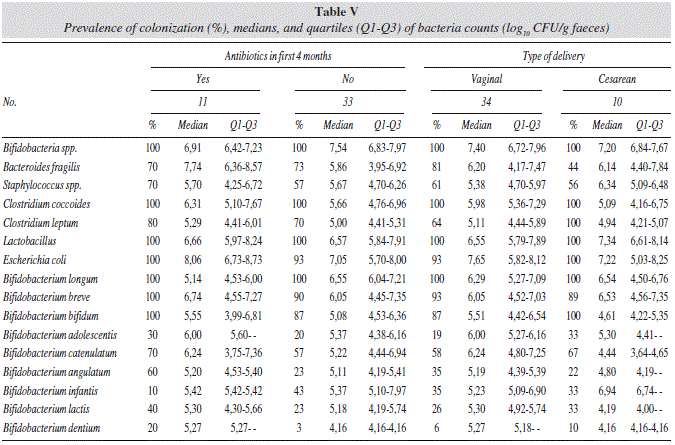
Infants whose mothers took antibiotic during pregnancy were associated with lower counts of B. angulatum.
Regarding milk-feeding practices, formula-feeding was associated with lower counts of B. angulatum in the infant.
Antibiotics intake in the first 4 months of life was associated with higher counts of Bacteroides fragilis group and B. angulatum, and lower counts of B. longum (table V). Spearman tests also showed significant correlations between infant's antibiotic intake and higher counts of Bacteroides fragilis group (R = 0.433; P = 0.019), higher B. angulatum prevalence (R=0.339; P = 0.032), and lower counts of Bifidobacterium spp. (R = -0.323; P = 0.042) and B longum (R = -0.443; P = 0.004).
Allergy and dermatitis incidence at follow-up was associated with lower counts of B. angulatum at 4 months.
Rotavirus vaccine administration was associated with lower counts of B. angulatum and was also significantly correlatedwith a lower prevalence of Bacteroides fragilis group (R = -0.396; P = 0.017). The rest of analysed bacteria did not show significant results for any of the EEF evaluated.
Finally, the logistic regression analysis did not show significant associations between the different EEF and the prevalence of bacterial colonization.
Discussion
Several EEF could be involved in immune system development and intestinal colonization in early life. It is well known that gut microbiota interacts with the gut-associated lymphoid tissue (GALT) and could thereafter modulate the systemic immune system. In this study, we assessed the associations between different EEF, peripheral lymphocyte subsets, and microbiota composition in fifty-five 4 month-old infants with at least one first-degree relative suffering from CD.
A widespread increase in the number of lymphocytes was associated with formula feeding in the studied population. The increase in lymphocytes involved mainly the T helper subsets, the overall subset (CD4+CD38+ and CD4+CD28+) as well as specifically the memory helper T cells. Similarly, an increase in the CD3+ and CD4+ lymphocyte counts were also associated with infant s infections, although in these cases, in addition to increased memory cell counts, also an increase in activated subsets expressing CD4+CD25+, and CD4+HLA-DR+ were associated with infection processes. Similar to our results, a study carried out with infants (0 to 6 months of age) of healthy mothers and uninfected infants of HIV-infected mothers, in which CD4+ lymphocyte phenotypes were examined, found that the proportions of activated (CD4+ HLA-DR+ CD38+) and memory (CD4+ CD45RA- RO+) lymphocytes were increased in uninfected infants of HIV-infected mothers compared to infants of uninfected mothers. Therefore, it was concluded that the lymphocytes of some HIV-exposed, uninfected infants, had been stimulated by antigen at early age and the cells principally implicated were helper T cells.26 Early infections would reasonably result in a higher proportion of memory and activated T helper cells as shown in our infants, however, regarding the association of formula-feeding with higher memory helper T cells, no similar study has been found in the literature to compare with, although an increase in T cells, particularly in T helper cells,15,17 and a decrease in NK cells17 have been reported for FF infants compared with BF infants at 6 months of age. Surprisingly, on the contrary to formula feeding and infant s infections, lower numbers of memory CD4+ cells were associated with the intake of antibiotics in the first 4mo., which suggest that taking antibiotics to halt the infections, alters the balance of immune cells acquired when infection is being eliminated by the body's own defense mechanisms. In vivo studies indicate that shortening the length of Listeria monocytogenes infection by antibiotic treatment can affect CD4+ T cell activation and development of memory T cells, but this effect is dependent on infectious dose, suggesting that overall antigen levels may play a role.27
The percentage of CD4+CD25+ cells was stimulated by vaginal delivery, antibiotics use by mothers during pregnancy, and breast-feeding, but in contrast, a lower percentage was associated with infants vaccinated against rotavirus. A recent study28 that compared the patterns of gene expression in peripheral blood mononuclear cells (PBMCs) from children with rotavirus diarrhea and healthy children showed that the first group had increased expression of genes involved in B-cell differentiation, maturation, activation and survival, but lower levels of mRNA for genes involved in the various stages of T-cell development. Importantly, this study also demonstrated a reduction in the total lymphocyte population and in the proportions of CD4+ and CD8+ T cells in PBMCs, suggesting that rotavirus alters T-cell homeostasis. According to our results, the rotavirus vaccine also influences the balance between T and B lymphocytes, as we have found by T test analysis that B cell percentage is higher in vaccinated infants (data not shown). On the other hand, previous results by our group in a bigger population with similar characteristics showed a higher percentage of CD4+CD25+ cells in BF infants than in FF infants,20 which is in agreement with the results in the current study.
According to the hygiene hypothesis, early infections caused by certain microorganisms are suspected to accelerate the Th1 maturation in children and to protect against future allergic disease.29 Our study showed that suffering infections during the first 4 months of life was associated with higher NK cell numbers. A similar result was found in a study comparing septic and healthy infants and children that described increased NK cells in the infants even after recovery from sepsis.30 Moreover the development of allergy or dermatitis within the first 18 months of life was associated with a lower percentage of NK cells at 4 months. Accordingly, studies in newborns at risk for allergy,31 infants with multiple allergy food,32 and children with atopic dermatitis 33 showed a decreased percentage32,33 and counts31 of NK cells. Our data suggest that infections suffered during the first 4 months of life might be protective, in the long term, for allergy and dermatitis development through the effect of infection on NK cells. However, no correlation was found between infection during the first 4 months and allergy incidence in the next months. Although, we have not found this correlation, the study by Han et al.34 support this last hypothesis. They suggest that NK cells may play an important role in infection-mediated inhibition of allergic responses. They studied the effect of chlamydial infection on the development of allergic responses induced by ovalbumin (OVA) and the involvement of NK cells in this process using a mouse model of airway inflammation. They found that the adoptive transfer of NK cells that were isolated from infected mice and injected to a syngeneic naive mice before OVA sensitization, produced a decrease of circulating and infiltrating eosinophils in the lung and inhibited the production of Th2 cytokines (IL-4 and IL-5) of spleen cells after sensitization.
Infants born by cesarean delivery are deprived of contact with their mother's intestinal and vaginal microbiota. In this infants, the acquisition of bacteroides , bifidobacteria, and E. coli has been shown to be delayed.8 Accordingly, we found that cesarean delivery was associated with lower counts of B. catenulatum. Similar results were also found in the KOALA study, which showed that infants born by cesarean section had lower numbers of bifidobacteria and bacteroides.35 On the other hand, a retrospective, multicenter, case-control study that included 1950 children found a significantly enhanced likelihood of being born by cesarean delivery in children with CD compared with control subjects.36 The authors hypothesized that the influence of cesarean delivery on the postnatal establishment of the intestinal microbiota might primarily affect the neonatal period and therefore enhance the intestinal epithelial permeability to gluten, leading to an aberrant stimulation of the mucosal immune system. This matter will possibly be elucidated after adequate length of follow-up is achieved for the cohort of the current study.
Oral use of antibiotics by the infant during the first 4 months also showed several associations with faecal microbiota composition, such as lower counts of B. longum and Bifidobacterium spp. The KOALA study also found lower counts of bifidobacteria in infants who received oral antibiotic therapy in their first 1 month.36 Finally, counts of B. angulatum appear to be associated with many of the different environmental factors studied (type of delivery, mother's antibiotic intake during pregnancy, milk-feeding practices, infections and antibiotic intake in the first 4 months of life, and rotavirus vaccine administration) which suggest that this might be a bacterial species more susceptible to fluctuations that others in its group.
One important point to consider about this study is the fact that it was performed in a population selected by the familial risk to suffer CD, which means that our results cannot be extrapolated to the general population. In this sense, we know that the frequency of infants with HLA haplotype different from DQ-2 in the group studied is significantly lower that the proportion in the general population.20 On the other hand, we have observed previously in a similar population that no interaction effect exists between milk-feeding practices and HLA-genotype on lymphocyte subsets in these infants.20 However, we cannot rule out the selection bias effect and therefore the importance of the influence of EEF on lymphocyte subsets and microbiota development would only be known in the future, when we are able to assess if the associations found here impact on CD development in this infant cohort.
In conclusion, infant s infections and antibiotic intake in the first 4 months of life are the EEF more strongly and/or frequently associated to lymphocyte subpopulations and microbiota composition, respectively in infants at risk of CD, but other factors such as milkfeeding practices, type of delivery, prenatal exposure to antibiotics, and rotavirus vaccine were also associated. The balance between the effects on lymphocyte subpopulations and microbiota composition of all these pre-and post-natal factors might modulate and define the risk for the future development of immune-related diseases such as CD. Thus, longer follow up of the entire PROFICEL cohort is necessary to assess the combined effect of environmental factors on that risk.
Acknowledgments
We gratefully acknowledge the assistance of the statistician Dr. Laura Barrios in the statistical analysis. T. Pozo-Rubio, G. De Palma and M. Olivares are recipient of a personal grant from the JAE Program of CSIC (Spain) cofunded by European Social Fund.
Conflict of interest statement
The authors declare that they have no conflict of interest.
References
1. Gianfrani C, Auricchio S, Troncone R. Adaptive and innate immune responses in celiac disease. Immunol Lett 2005; 99: 141-5. [ Links ]
2. Meresse B, Ripoche J, Heyman M, Cerf-Bensussan N. Celiac disease: from oral tolerance to intestinal inflammation, autoimmunity and lymphomagenesis. Mucosal Immunol 2009; 2:8-23. [ Links ]
3. Dubois PC, van Heel DA. Translational mini-review series on the immunogenetics of gut disease: immunogenetics of coeliac disease. Clin Exp Immunol 2008; 153: 162-73. [ Links ]
4. Nova E, Pozo T, Sanz Y, Marcos A. Dietary strategies of immunomodulation in infants at risk for celiac disease. The Proceedings of the Nutrition Society 2010; 69: 347-53. [ Links ]
5. Westerbeek EA, van den Berg A, Lafeber HN, Knol J, Fetter WP, van Elburg RM. The intestinal bacterial colonisation in preterm infants: a review of the literature. Clin Nutr 2006; 25: 361-8. [ Links ]
6. Williams AM, Bland PW, Phillips AC, Turner S, Brooklyn T, Shaya G, et al. Intestinal alpha beta T cells differentiate and rearrange antigen receptor genes in situ in the human infant. J Immunol 2004; 173: 7190-9. [ Links ]
7. McLoughlin RM, Mills KH. Influence of gastrointestinal commensal bacteria on the immune responses that mediate allergy and asthma. The Journal of Allergy and Clinical Immunology 2011; 127: 1097-107; quiz 108-9. [ Links ]
8. Adlerberth I, Wold AE. Establishment of the gut microbiota in Western infants. Acta Paediatrica 2009; 98: 229-38. [ Links ]
9. De Vries E, de Bruin-Versteeg S, Comans-Bitter WM, de Groot R, Hop WC, Boerma GJ, et al. Longitudinal survey of lymphocyte subpopulations in the first year of life. Pediatr Res 2000; 47: 528-37. [ Links ]
10. Comans-Bitter WM, de Groot R, van den Beemd R, Neijens HJ, Hop WC, Groeneveld K, et al. Immunophenotyping of blood lymphocytes in childhood. Reference values for lymphocyte subpopulations. J Pediatr 1997; 130: 388-93. [ Links ]
11. Panaro A, Amati A, di Loreto M, Felle R, Ferrante M, Papadia AM, et al. Lymphocyte subpopulations in pediatric age. Definition of reference values by flow cytometry. Allergol Immunopathol (Madr) 1991; 19: 109-12. [ Links ]
12. Shearer WT, Rosenblatt HM, Gelman RS, Oyomopito R, Plaeger S, Stiehm ER, et al. Lymphocyte subsets in healthy children from birth through 18 years of age: the Pediatric AIDS Clinical Trials Group P1009 study. J Allergy Clin Immunol 2003; 112: 973-80. [ Links ]
13. O'Gorman MR, Millard DD, Lowder JN, Yogev R. Lymphocyte subpopulations in healthy 1-3-day-old infants. Cytometry 1998; 34: 235-41. [ Links ]
14. McCloskey TW, Cavaliere T, Bakshi S, Harper R, Fagin J, Kohn N, et al. Immunophenotyping of T lymphocytes by three-color flow cytometry in healthy newborns, children, and adults. Clinical Immunology and Immunopathology 1997; 84: 46-55. [ Links ]
15. Carver JD, Pimentel B, Wiener DA, Lowell NE, Barness LA. Infant feeding effects on flow cytometric analysis of blood. J Clin Lab Anal 1991; 5: 54-6. [ Links ]
16. Cummins AG, Eglinton BA, Gonzalez A, Roberton DM. Immune activation during infancy in healthy humans. J Clin Immunol 1994;14:107-15. [ Links ]
17. Hawkes JS, Neumann MA, Gibson RA. The effect of breast feeding on lymphocyte subpopulations in healthy term infants at 6 months of age. Pediatr Res 1999; 45: 648-51. [ Links ]
18. Jeppesen DL, Hasselbalch H, Lisse IM, Ersboll AK, Engelmann MD. T-lymphocyte subsets, thymic size and breastfeeding in infancy. Pediatr Allergy Immunol 2004; 15: 127-32. [ Links ]
19. Andersson Y, Hammarstrom ML, Lonnerdal B, Graverholt G, Falt H, Hernell O. Formula feeding skews immune cell composition toward adaptive immunity compared to breastfeeding. Journal of immunology 2009; 183: 4322-8. [ Links ]
20. Pozo-Rubio T, Capilla A, Mujico IR, de Palma G, Marcos A, Sanz Y, et al. Influence of breastfeeding versus formula feeding on lymphocyte subsets in infants at risk of coeliac disease: the PROFICEL study. Eur J Nutr 2012. [ Links ]
21. Duijts L, Bakker-Ionges LE, Labout IA, laddoe VW, Hofman A, Steegers EA et al. Perinatal stress influences lymphocyte subset counts in neonates. The generation R study. Pediatric Research 2008; 63: 292-8. [ Links ]
22. Samelson R, Larkey DM, Amankwah KS, McConnachie P. Effect of labor on lymphocyte subsets in full-term neonates. Am J Reprod Immunol 1992; 28: 71-3. [ Links ]
23. Chirico G, Gasparoni A, Ciardelli L, Martinotti L, Rondini G. Leukocyte counts in relation to the method of delivery during the first five days of life. Biol Neonate 1999; 75: 294-9. [ Links ]
24. Matsuki T, Watanabe K, Fujimoto I, Miyamoto Y, Takada T, Matsumoto K, et al. Development of 16S rRNA-gene-targeted group-specific primers for the detection and identification of predominant bacteria in human feces. Applied and Environmental Microbiology 2002; 68: 5445-51. [ Links ]
25. Malinen E, Kassinen A, Rinttila T, Palva A. Comparison of real-time PCR with SYBR Green I or 5'-nuclease assays and dot-blot hybridization with rDNA-targeted oligonucleotide probes in quantification of selected faecal bacteria. Microbiology 2003; 149: 269-77. [ Links ]
26. Rich KC, Siegel IN, lennings C, Rydman RI, Landay AL. Function and phenotype of immature CD4+ lymphocytes in healthy infants and early lymphocyte activation in uninfected infants of human immunodeficiency virus-infected mothers. Clinical and Diagnostic Laboratory Immunology 1997; 4: 358-61. [ Links ]
27. Blair DA, Lefrancois L. Increased competition for antigen during priming negatively impacts the generation of memory CD4 T cells. Proceedings of the National Academy of Sciences of the United States of America 2007; 104: 15045-50. [ Links ]
28. Wang Y, Dennehy PH, Keyserling HL, Tang K, Gentsch IR, Glass RI, et al. Rotavirus infection alters peripheral T-cell homeostasis in children with acute diarrhea. Journal of virology 2007; 81: 3904-12. [ Links ]
29. Blanco Quiros A, Arranz Sanz E. Early infections and later allergic diseases. Allergologia et Immunopathologia 2009; 37:279-80. [ Links ]
30. Wisniewska-Ligier M, Wozniakowska-Gesicka T, Zeman I, Banasik M. Evaluation of lymphocyte subsets and NK cells in septic children. Med Sci Monit 2002; 8: CR119-24. [ Links ]
31. Liao SY, Liao TN, Chiang BL, Huang MS, Chen CC, Chou CC, et al. Decreased production of IFN gamma and increased production of IL-6 by cord blood mononuclear cells of newborns with a high risk of allergy. Clinical and experimental allergy: Journal of the British Society for Allergy and Clinical Immunology 1996; 26: 397-405. [ Links ]
32. Latcham F, Merino F, Lang A, Garvey I, Thomson MA, Walker-Smith IA, et al. A consistent pattern of minor immunodeficiency and subtle enteropathy in children with multiple food allergy. The Journal of Pediatrics 2003; 143: 39-47. [ Links ]
33. Campbell DE, Fryga AS, Bol S, Kemp AS. Intracellular interferon-gamma (IFN-gamma) production in normal children and children with atopic dermatitis. Clinical and Experimental Immunology 1999; 115: 377-82. [ Links ]
34. Han X, Fan Y, Wang S, Iiao L, Qiu H, Yang X. NK cells contribute to intracellular bacterial infection-mediated inhibition of allergic responses. Journal of Immunology 2008; 180: 4621-8. [ Links ]
35. Penders I, Thijs C, Vink C, Stelma FF, Snijders B, Kummeling I, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics 2006; 118: 511-21. [ Links ]
36. Decker E, Engelmann G, Findeisen A, Gerner P, Laass M, Ney D, et al. Cesarean delivery is associated with celiac disease but not inflammatory bowel disease in children. Pediatrics 2010; 125: e1433-40. [ Links ]
![]() Correspondence:
Correspondence:
Tamara Pozo-Rubio.
Immunonutrition Group, Department of Metabolism and Nutrition.
Institute of Food Science, Technology and Nutrition (ICTAN).
Spanish National Research Council (CSIC).
C/ José Antonio Novais 10.
28040 Madrid. Spain.
E-mail: tpozo@ictan.csic.es
Recibido: 10-XI-2012.
Aceptado: 12-XII-2012.

