










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkNutrición Hospitalaria
versión On-line ISSN 1699-5198versión impresa ISSN 0212-1611
Nutr. Hosp. vol.33 no.1 Madrid ene./feb. 2016
https://dx.doi.org/10.20960/nh.39
TRABAJO ORIGINAL / Otros
Protection by polyphenol extract from olive stones against apoptosis produced by oxidative stress in human neuroblastoma cells
Protección de un extracto de polifenoles de huesos de aceitunas frente a la apoptosis producida por estrés oxidativo en células de neuroblastoma humano
Ernesto Cortés-Castell1, Carmen Veciana-Galindo2, Luis Torró-Montell2, Antonio Palazón-Bru3, Elia Sirvent-Segura2, Vicente Gil-Guillén3 and Mercedes Rizo-Baeza4
1Department of Pharmacology, Pediatrics and Organic Chemistry. Universidad Miguel Hernández. Alicante.
2Laboratorio de Biotecnología y Proyectos. Biopartner S.L. Alicante.
3Department of Clinical Medicine. Universidad Miguel Hernández. Alicante.
4Department of Nursing and Nutrition. Universidad de Alicante. Alicante
ABSTRACT
Objective: We evaluated the protective activity of an extract from a by-product such as olive stones, through its ability to inhibit H2O2 induced apoptosis in the SH-SY5Y human neuroblastoma cell line.
Material and methods: To such end, 20,000 cells/well were cultivated and differentiation with retinoic acid was initiated. Once the cells were differentiated, apoptosis was induced with and without H2O2 extract. Finally, cDNA extraction was performed, and pro-apoptotic genes Bax and anti-apoptotic genes Bcl-2 were analyzed. Quantification of the gene expression was performed using the GAPDH gene marker.
Results: Cell viability with the extract is 97.6% (SD 5.7) with 10 mg/l and 62.8% (SD 1.2) to 50 mg/l, using 10 mg/l for the biomarker assay. The retinoic acid differentiated SH-S cell line (10 µM) shows a clear apoptosis when treated with H2O2 150 µM, with a Bax/Bcl-2 ratio of 3.75 (SD 0.80) in contrast to the differentiated control cells subjected to H2O2 and with extract, which have the same ratio of 1.02 (SD 0.01-0.03).
Conclusion: The olive stone extract shows anti-apoptotic activity in the provoked cell death of SH-SY5Y human neuroblastoma cells in their normal state, defending them from oxidative stress which produces a significant increase in the apoptotic gene ratio in contrast to anti-apoptotic genes (Bax/Bcl-2).
Key words: Polyphenols. Olive stones. Apoptosis. Human neuroblastome cells.
RESUMEN
Objetivo: evaluar la actividad protectora del extracto de un subproducto como son los huesos de aceitunas, mediante su capacidad de inhibir la apoptosis en la línea celular humana de neuroblastoma SH-SY5Y inducida con H2O2.
Material y métodos: se han cultivado 20.000 cel/pocillo, iniciando diferenciación con ácido retinoico y, una vez diferenciadas las células, se ha inducido la apoptosis con H2O2 con extracto y sin presencia del mismo. Finalmente se efectúa la extracción de cDNA y el análisis de los genes proapoptótico Bax y antiapoptótico Bcl-2. La cuantificación de la expresión génica se realiza frente al gen marcador GAPDH.
Resultados: la viabilidad celular con el extracto es del 97,6% (SD 5,7) con 10 mg/l y 62,8% (SD 1,2) a 50 mg/l, utilizando 10 mg/l para el ensayo de biomarcadores. Las células de la línea SH-S diferenciadas con ácido retinoico (10 µM), muestran una clara apoptosis al ser tratadas con H2O2 150 µM, con una relación Bax/Bcl2 de 3,75 (SD 0,80) frente a las células diferenciadas control y sometidas a H2O2 y con extracto que tienen la misma relación de 1,02 (SD 0,01-0,03).
Conclusión: el extracto de huesos de aceitunas presenta una actividad antiapoptótica frente a la provocación de la muerte celular por peróxido de hidrógeno, preservando a las células de neuroblastoma humano SH-SY5Y en su estado de normalidad, al defenderlas del estrés oxidativo que produce un significativo aumento de la relación de genes apoptóticos frente a antiapoptóticos (Bax/Bcl2).
Palabras clave: Polifenoles. Huesos de aceitunas. Apoptosis. Células de neuroblastoma.
Introduction
Antioxidants can prevent the harmful effects of free radicals in cells in the same way that a diet rich in polyphenols can decrease the risk of cancer (1), heart disease, and some neurological diseases (2), and prevent neurodegeneration caused by oxidative stress, among others (3).
Olive trees constitute a natural source of bioactive compounds. The processing of olives leads to different types of secondary matrices, which are generally considered as by-products. One characteristic these by-products have in common with plant waste generated from agroalimentary activity is their potential as a source of bioactive compounds. Therefore, a way of evaluating these residues is to use them as a raw material for extracting compounds with satiating properties: fiber, fatty acids and proteins, which have a high economic value and can be used for their satiating and neuroprotective properties in the agroalimentary and pharmaceutical industry, or even as functional compounds in foods. One area of focus regarding olive trees is to obtain aqueous effluents from the olive oil process (4). In other cases, the recuperation of substances of interest focuses on other types of matrices like solid waste: pulp, leaf or olive wood. The major component of olive leaf is oleuropein, it is also abundant in hydroxytyrosol (5), which is the main compound derived from oleuropein and has high antioxidant ability. However, the general activity from these extracts seems to be supported by the synergetic effects between the substances they are composed of. The polyphenols from olive trees have become the subject of many studies due to their powerful antioxidant activity and anti-inflammatory properties.
The importance of phenolic compounds in diets is mainly due to their antioxidant ability. They are scavengers of free radicals and have the ability to chelate transition metal ions, thereby stopping oxidative chain reactions in cells (6). All the polyphenols studied show a high antioxidant activity (7). The most abundant phenolic compound in olive tree extracts is (3.4-dihydroxyphenyl) ethanol or hydroxytyrosol: it presents scavenging activity of the superoxide radical generated by the hypoxanthina-xanthine oxidase system, and of the hydrogen peroxide generated by the reaction of guaiacol (8); it inhibits oxidation of salicylic acid by the radical -OH9; it reduces the catalase inactivation mediated by hypochlorous acid; it can act on reactive species of nitrogen like peroxynitrite (8); and it acts against linoleic acid autooxidation in lipophilic environments, as a metal chelantor -for example, iron and copper, which are essential for the organism, but can be potentially dangerous since they are capable of transferring electrons and acting as catalyzers of autooxidation reactions.
These antioxidant properties suggest that hydroxytyrosol and other polyphenols (verbacoside, elenolic acid, cycloolivil, caffeic acid, gallic acid, etc.) that are present in the olive tree could have a protector role against the oxidation of lipoproteins and in this way they can contribute to the prevention of cardiovascular disease (9). Its anti-cancer properties are also apparent in the context that the generation of free radicals seem to be involved in cancer pathology, suggesting that these phenolic compounds can be related to the low incidence of different types of cancer associated with diets which are rich in these compounds (10), by protecting the DNA of prostate cells from harm caused by free radicals, thereby preventing mutagenic activity caused by oxidative stress (11) and by ultraviolet light (12).
These polyphenols also inhibit platelet aggregation (13) and present in vitro antimicrobial properties against several infectious agents from the gastrointestinal and respiratory tract (14). They have also been attributed a neuroprotector function (8,15), digestive protection, as well as an antihypercholesterolemic and antihyperglycemic regulator of adipose tissue.
Although other studies have shown various biochemical characteristics (16,17), no one has determined the neurological protection effectiveness of the olive seed extract. Therefore, we performed a study evaluating the anti-apoptotic activity of an extract from olive stones, rich in polyphenols, in cell death induced by oxidative stress in human neuroblastoma cultures.
Material and methods
The study of the activity of the olive stone extracts was carried out through the study of protection against apoptosis, using SH-SY5Y human neuroblastoma cell line (94030304, ECCC) (18,19). To do so, "neuronal death", apoptosis, was induced in vitro and an evaluation was made of how the presence of the extracts affects the process, analyzing two marker genes: the Bax gene from the pro-apoptotic gene family ("cell death genes") and the Bcl-2 gene from the anti-apoptotic gene family ("survival genes"). The neuroprotector effect was considered as such if the ratio between both molecules decreased: Bax/Bcl-220.
To perform the assay, cells were seeded into 24-well plates at a concentration of 20,000 cell/ well and were incubated at 37 oC and 5% CO2 in the medium recommended by ECACC (Ham's F12:EMEM [EBSS] [1:1] + 2mM L-Glutamine + 1% Non-Essential Amino Acids [NEAA] + 15% Fetal Bovine Serum [FBS]). After 24 hours incubation, differentiation was started with retinoic acid 10 μM (Sigma transretinoic acid [RA]) in a medium with 1% FCS for 6 days, changing the medium every three days. One day before the experiment, the medium was replaced with specific medium without serum and 1% N2 (Invitrogen) supplement. After 24 hours incubation, the extract treatment was started at the selected and previously described biosecure concentrations (21). They were incubated for 2 hours with the extracts (10 and 50 mg/l), cell viability was measured, and afterwards cell damage was induced with H2O2 150 μM for 5 hours (22,23). Once the treatment was finished the cells were collected for rt-PCR molecular analysis.
The analysis of the gene expressions was made from the cells by means of RNA extraction. The extraction was performed with the Rneasy Mini Kit (74104 QIAGEN). c-DNA was obtained from the RNA through the transcriptase enzyme (High Capacity cDNA reverse transcription kit [4368814] Applied Biosystems). PCR was performed in real time from the cDNA, so for the pro-apoptotic BAX gene the primers Hs00180269_m1 from Applied Biosystems were used; and for the anti-apoptotic Bcl-2 gene the primers Hs00608023_m1 from Applied Biosystems were used, and GAPDH (Primers Hs02758991_g1) were used as control gene. The reaction conditions were 10 μl (1X) TaqMan Gene Expression Mastermix 4369016 Applied Biosystems; 2 μl (0.3 μM) of each primer, 2 μl (0.1μM) of gene marker serum, 0.4 μl of PCR internal control, (DNA exo 50X), 2 μl Mix EXO and 2 μl (50 ng) of c-DNA.
The amplification conditions in the thermocycler (7300 Applied Biosystem) were universal: 50 oC for 2 min, 95 oC for 10 min and 40 cycles of: 95 oC-15 sec and 60 oC-1 min.
Relative quantification of genetic expression was carried out, obtaining the scale of the physiological changes in the biomarker gene in comparison with a reference gene. The formula 2-ΔΔCt was used for the calculations.
Quantification was measured in each PCR cycle by the amount of amplicon produced through the addition of the fluorophores that become attached to the amplicon quantitatively. The real time PCR system detects the quantity of fluorescence produced in each PCR cycle and the software analysis represents this fluorescence graphically in relation to the number of cycles. The quantity of amplicon produced is proportional to the number of initial RNA/DNA molecules, so that in those samples with a greater gene expression, fluorescent amplicon will appear in the previous cycles. Reference samples were the live cells (without cell damage) and the GAPDH gene was the internal control gene.
All the assays were carried out in two different periods and duplicated, so the result is the average of four values. The statistical study was made by calculating averages, standard deviations and intervals of confidence at 95% (95% CI), as well as the comparison between groups (t-test) using the Epidat 3.1 software.
Results and discussion
The viability of the SH-SY5Y human neuroblastoma cell cultures, prior to the cell damage induced by hydrogen peroxide, is 97.6% (SD 5.7) (95% CI: 88.5-100%) at 10 mg/l of extract, and 62.8% (SD 1.2) (95% CI: 60.9-64.7%) at 50 mg/l. As viability was lower than 80% (biosecurity limit) at 50 mg/l concentration, only the 10 mg/l concentration of extract was used for the genetic biomarkers assay.
Figure 1 shows the SH-S cell line differentiated with retinoic acid (10 µM), before and after being treated with H2O2 150 µM.

The results for the genetic biomarkers at 10 mg/l are shown in table I, where the ratio of the Bax and Bcl-2 gene expression is shown. The data given is related to the control cells (without oxygenated water treatment). In cell damage conditions, the expression of the Bax and Bcl-2 ratio increases indicating higher pro-apoptotic gene (Bax) ratio in contrast to anti-apoptotic (Bcl-2) gene. This occurs when the cell culture is subjected to H2O2 150 μM and, therefore, shows a tendency towards early cell death (apoptosis). For cell viability of the extract (10 mg/l), although the culture is subjected to hydrogen peroxide, which is an agent that clearly induces cell death, it is possible to observe a balanced ratio of gene expression, which is exactly the same as the one that occurs without the cell death inductor and the extract.
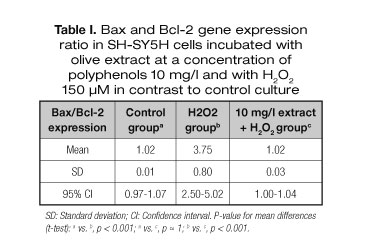
Therefore, the extract has an anti-apoptotic ability on oxidative stress in human neuroblastoma cells, which equals the ratio of pro-apoptotic and anti-apoptotic genes in their normal state. If 100% cell viability is considered as normal conditions, the effect of the extract presents a cytoprotector effect of 100%.
To date, neurodegenerative pathologies of multifactorial origin present a difficult scenario for establishing therapeutic strategies. Nevertheless, neuroprotector mechanisms can be dealt with from dietary modifications. The polyphenols contained in olive tree extracts present antioxidant properties that make them candidates for research into neurodegenerative diseases (24). Apart from other beneficial effects, the protective power against dementia from the intake of flavonoids and phenolic compounds contained in wines, vegetables and fruit has been demonstrated (25).
In vitro and in vivo evaluation of bioactive compounds is based on experimental models that reproduce the biological process of interest through systems which are sensitive to external factors and whose possible fluctuations are measurable. The global objective of this paper was the study of the neuroprotector ability from a natural compound obtained from the olive stones applied to in vitro models in cell lines. To do so, the SH-SY5Y human neuroblastoma cell line (94030304, ECCC) was used (18,19), whose differentiation leads to cells which are similar to human neurons. The result shows some cells that present a postmitotic neuronal phenotype with a well-defined morphology and polarity, and the expression of typical neuronal markers (26). These SH-SY5Y cells constitute a homogenous and highly reproducible model of cells which are similar to human neurons in culture. Nowadays, a multitude of apoptotic stimulants are used in order to study the molecular mechanisms which lead to cell death, for example: the withdrawal of trophic factors, oxygen deprivation, thermic shock, or the administration of toxic agents which selectively cause harm in DNA or in certain cell organs (27). For the evaluation of the effect of the extract on apoptosis, gene markers were analyzed: the Bax gene from the pro-apoptotic family ("death cell genes") and the Bcl-2 gene from the anti-apoptotic family ("survival gens"), thus the balance between both defines the threshold of programmed cell death (28,29). The neuroprotector effect is considered as such, if the ratio between both molecules Bax/Bcl-2 decreases (20). This study has shown the neuroprotector effect of olive stone extract on apoptosis induced by hydrogen peroxide.
Other authors have found similar effects using different models, like Schaffer et al. (30) who use the PC12 cell line from brain cells to study polyphenols from waters from the olive process. They subjected the cells to oxidative stress and measured cytotoxicity, observing a cytoprotection of the cerebral cells with extracts. Other authors have used primary cultures from mouse neurons and mixed culture systems of cerebellar cells enriched by Purkinje neurons (31). These neurons, which constitute the only efferent projection of the cerebellum, present a low rate of survival in culture, but constitute a very interesting model for the study of neuron-glia interactions (32) and other neuronal functions (33).
A possible mechanism of protection against apoptosis to be described is the inhibition of the glycogen synthase kinase enzyme - 3(GSK-3), which provokes a protector effect against pro-apoptotic stimulants (34,35). So, after verifying that GSK-3 inhibition increases the anaerobic intake of glucose through glycolysis, measured by extracellular release of lactate, this metabolic change was analyzed to see whether it is at least partly responsible for the neuroprotector effect against death by mitochondrial dysfunction. By inhibiting the glycolytic pathway with 2-desoxiglucose (2-DGlc), it can be observed that it completely neutralizes the neuroprotector effect of chronic inhibition from GSK-3, supporting the hypothesis of a change towards the glycolytic metabolism which is less dependent on mitochondrial respiration.
Another possible neuroprotector mechanism is the possible involvement of neurotrophin BDNF (neurtrophic factor), given its role in anti-apoptotic signaling, as well as its neuroprotector effect in some neuronal damage models (36,37).
Recent studies suggest that olive tree extracts inhibit inflammation (38) and reduce oxidative stress, which has been observed in rats with a provoked cerebral isquemia (38). For example, the neuroprotector effects of dry olive leave extract on global transient cerebral isquemia in Mongolian gerbils (39), and the evaluation of different parameters of oxidative stress and neuronal damage in the hippocampus. These effects were compared to those of quercatin, which is a flavonoid known to be a neuroprotector. The treatment with this extract significantly inhibits the production of superoxide and nitric oxide, decreases lipid peroxidation and increases the activity of the superoxide dismutase, the effects being significantly higher than those from quercatin, indicating that it exercises a potent neuroprotector activity against neuronal damage in the hippocampus after global transient cerebral isquemia, which can be attributed to its antioxidant properties. The majority of studies about the effects of polyphenols from olive tree extracts indicate that their neuroprotector action is due to their antioxidant ability by eliminating reactive oxygen and nitrogen species involved in human diseases (8).
Conclusions
The olive stone extract presents an anti-apoptotic activity against the provocation of cell death by hydrogen peroxide, returning the SH-SY5Y human neuroblastoma cells to their normal state, by defending them against oxidative stress which produces a significant increase in the apoptotic gene ratio in contrast to the anti-apoptotic gene (Bax/Bcl-2).
Acknowledgments
The authors thank Catherine Nicholls Baeza for help with the English language version of the text.
Conflicts of interest
Laboratorio de Biotecnología y Proyectos, Biopartner S.L. (Alcoy, Spain) is performing the patent process for the extract carried out in this work.
References
1. Tangney CC, Rasmussen HE. Polyphenols, inflammation, and cardiovascular disease. Curr Atheroscler Rep 2013;15:324. [ Links ]
2. Bartlett H, Eperjesi F. Age-related macular degeneration and nutritional supplementation: a review of randomised controlled trials. Ophthalmic Physiol Opt 2003;23:383-99. [ Links ]
3. Bleys J, Miller E, Pastor-Barriuso R, Appel L, Guallar E. Vitamin-mineral supplementation and the progression of atherosclerosis: a meta-analysis of randomized controlled trials. Am J Clin Nutr 2006;84:880-7. [ Links ]
4. De Marco E, Savarese M, Paduano A, Sacchi R. Characterization and fractionation of phenolic compounds extracted from olive oil mill wastewaters. Food Chem 2007;104:858-67. [ Links ]
5. Pereira-Caro G, Madrona A, Mateos R, Rodríguez G, Trujillo M, et al. Synthesis of hydroxytyrosyl alkyl ethers from olive oil waste waters. Molecules 2009;14:1762-72. [ Links ]
6. Halliwell B, Rafter J, Jenner A. Health promotion by flavonoids, tocopherols, tocotrienols, and other phenols: direct or indirect effects? Antioxidant or not? Am J Clin Nutr 2005;81:268S-276S. [ Links ]
7. Obied HK, Prenzler PD, Konczak I, Rehman AU, Robards K. Chemistry and bioactivity of olive biophenols in some antioxidant and antiproliferative in vitro bioassays. Chem Res Toxicol 2009;22:227-34. [ Links ]
8. De-la-Puerta R, Martinez-Dominguez ME, Ruiz-Gutierrez V. Effects of virgin olive oil phenolics on scavenging of reactive nitrogen species and upon nitergic neurotransmission. Life Sci 2001;69:1213-22. [ Links ]
9. Covas MI, Fito M, Lamuela-Raventos RM, Sebastia N, De-la-Torre-Boronat C, et al. Virgin olive oil poliphenolic compounds: binding to human low density lipoprotein (LDL) and effect on LDL oxidation. Int J Pharm Res 2000;20:49-54. [ Links ]
10. Owen RW, Haubner R, Wurtele G, Hull E, Spiegelhader B, et al. Olives and olive oil in cancer prevention. Eur J Cancer Prev 2004;13:319-26. [ Links ]
11. Quiles JL, Huertas JR, Ochoa JJ, Battino M, Mataix J, et al. Dietary fat (virgin olive oil or sunflower oil) and physical training interactions on blood lipids in the rat. Nutr 2003;19:363-8. [ Links ]
12. D'Angelo S, Ingrosso D, Migiarrdi V, Sorrentino A, Donnarumma G, et al. Hydroxytyrosol, a natural antioxidant from olive oil, prevents protein damage induced by long-wave ultraviolet radiation in melanoma cells. Free Radic Biol Med 2005;38:908-19. [ Links ]
13. Petroni A, Blasevich M, Salami M, Papini N, Montedoro GF, et al. Inhibition of platelet aggregation and eicosanoid production by phenolic components of olive oil. Thromb Res 1995;78:151-60. [ Links ]
14. Furneri PM, Piperno A, Sajia A, Bisignano G. Antimycoplasmas activity of hydroxytyrosol. Antimicrob Agents Chemother 2004;48:4892-4. [ Links ]
15. Vauzour D, Corona G, Spencer JP. Caffeic acid, tyrosol and p-coumaric acid are potent inhibitors of 5-S-cysteinyl-dopamine induced neurotoxicity. Arch Biochem Biophys 2010;501:106-11. [ Links ]
16. Cortés E, Veciana C, Torro L, Sirvent E, Rizo MM, et al. Actividad antiinflamatoria de un extracto polifenólico de hueso de olivas en la línea celular de monocitos humanos THP1-XBLUE-CD14. Nutr Hosp 2014;30:113-7. [ Links ]
17. Cortés E, Veciana C, Torro L, Sirvent E, Rizo MM, et al. Efecto sobre el neurodesarrollo y neuroprotección en pez cebra de un extracto polifenólico de huesos de aceituna. Nutr Hosp 2014;30:338-42. [ Links ]
18. Cheung YT, Lau WKW, Yu MS, Lai CSW, Yeung SC, et al. Effects of all-trans-retinoic acid on human SH-SY5Y neuroblastoma as in vitro model in neurotoxicity research. Neuro Toxicol 2009;30:127-35. [ Links ]
19. Zhang L, Yu H, Sun Y, Lin X, Chen B, et al. Protective effects of salidroside on hydrogen peroxide-induced apoptosis in SH-SY5Y human neuroblastoma cells. Eur J Pharmacol 2007;564:18-25. [ Links ]
20. Gao M, Zhang WC, Liu QS, Hu JJ, Liu GT, et al. Pinocembrin prevents glutamate-induced apoptosis in SH-SY5Y neuronal cells via decrease of bax/bcl-2 ratio. Eur J Pharmacol 2008;591:73-9. [ Links ]
21. Veciana C, Cortés E, Torro L, Sirvent E, Rizo M, et al. Evaluación de la citotoxicidad y bioseguridad de un extracto de polifenoles de huesos de aceitunas. Nutr Hosp 2014;29:1388-93. [ Links ]
22. Kim HJ, Song JY, Park HJ, Park HK, Yun DH, et al. Naringin Protects against Rotenone-induced Apoptosis in Human Neuroblastoma SHSY5Y Cells. Kor J Physiol Pharmacol 2009;13:281-5. [ Links ]
23. Jaworska-Feil L, Jantas D, Leskiewicz M, Budziszewska B, Kubera M, et al. Protective effects of TRH and its analogues against various cytotoxic agents in retinoic acid (RA)-differentiated human neuroblastoma SH-SY5Y cells. Neuropeptides 2010;44:495-508. [ Links ]
24. Sears B, Ricordi C. Role of fatty acids and polyphenols in inflammatory gene transcription and their impact on obesity, metabolic syndrome and diabetes. Eur Rev Med Pharmacol Sci 2012;16:1137-54. [ Links ]
25. Commenges D. Intake of flavonoids and risk of dementia. Eur J Epidemiol 2000;16:357-63. [ Links ]
26. Gimenez-Cassina A, Lim F, Diaz-Nido J. Differentiation of a human neuroblastoma into neuron-like cells increases their susceptibility to transduction by herpesviral vectors. J Neurosci Res 2006;84:755-67. [ Links ]
27. Lindahl PE, Oberg KE. The effect of rotenone on respiration and its point of attack. Exp Cell Res 1961;23:228-37. [ Links ]
28. Oltavi ZN, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell 1993;74:609-19. [ Links ]
29. Adams JM, Cory S. The Bcl-2 protein family: arbiters of cell survival. Science 1998;281:1322-6. [ Links ]
30. Schaffer S, Müller WE, Eckert GP. Cytoprotective effects of olive mill wastewater extract and its main constituent hydroxytyrosol in PC12 cells. Pharmacol Res 2010;62:322-7. [ Links ]
31. Agudo M, Trejo JL, Lim F, Avila J, Torres-Aleman I, et al. Highly efficient and specific gene transfer to Purkinje cells in vivo using a herpes simplex virus I amplicon. Hum Gene Ther 2002;13:665-74. [ Links ]
32. Morrison ME, Mason CA. Granule neuron regulation of Purkinje cell development: striking a balance between neurotrophin and glutamate signaling. J Neurosci 1998;18:3563-73. [ Links ]
33. Bravin M, Morando L, Vercelli A, Rossi F, Strata P. Control of spine formation by electrical activity in the adult rat cerebellum. Proc Natl Acad Sci USA 1999;96:1704-9. [ Links ]
34. Bijur GN, Jope RS. Glycogen synthase kinase-3beta is highly activated in nuclei and mitochondria. Neuroreport 2003;14:2415-9. [ Links ]
35. Linseman DA, Butts BD, Precht TA, Phelps RA, Le SS, et al. Glycogen synthase kinase-3beta phosphorylates Bax and promotes its mitochondrial localization during neuronal apoptosis. J Neurosci 2004;24:9993-10002. [ Links ]
36. Chao MV. Neurotrophins and their receptors: a convergence point for many signalling pathways. Nat Rev Neurosci 2003;4:299-309. [ Links ]
37. Perez-Navarro E, Gavalda N, Gratacos E, Alberch J. Brain-derived neurotrophic factor prevents changes in Bcl-2 family members and caspase-3 activation induced by excitotoxicity in the striatum. J Neurochem 2005;92:678-91. [ Links ]
38. Mohagheghi F, Bigdeli MR, Rasoulian B, Hashemi P, Pour MR. The neuroprotective effect of olive leaf extract is related to improved blood-brain barrier permeability and brain edema in rat with experimental focal cerebral ischemia. Phytomedicine 2011;18:170-5. [ Links ]
39. Dekanski D, Selakovi V, Piperski V, Radulovi Z, Koreni A, et al. Protective effect of olive leaf extract on hippocampal injury induced by transient global cerebral ischemia and reperfusion in Mongolian gerbils. Phytomedicine 2011;18:1137-43. [ Links ]
![]() Correspondence:
Correspondence:
Ernesto Cortés Castell.
Department of Pharmacology, Pediatrics and Organic Chemistry.
Universidad Miguel Hernández.
Avda. de la Universidad, s/n.
03202 Elche, Alicante
e-mail: ernestocort@gmail.com
Received: 07/10/15
Accepted: 15/11/15
