










 Servicios personalizados
Servicios personalizados
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCTION
Various studies have indicated an association between modification in dietary macronutrient composition and liver apoptosis, but the exact mechanism in the development of apoptosis under these circumstances is not entirely understood 1-4.
Apoptosis is an innately programmed process responsible for the death of unwanted cells. It occurs mainly during embryogenesis or in tissues with the capacity for regeneration such as the liver 5. Senescent and mutagenic cells that have completed their mission are eliminated by apoptosis, and new cells are formed when necessary to replace them 6. Dysregulation occurs when the regenerative capacity is lower than the apoptosis rate, and the physiological pathway becomes pathological.
Apoptosis leads to the development of numerous diseases such as cancer, autoimmune diseases, viral or bacterial infections, alcoholic liver disease, viral hepatitis, cholestatic liver disease, hepatocellular carcinoma, Wilson's disease, and nonalcoholic steatohepatitis 5.
There are two forms of triggered apoptosis. The first is mediated by agents that penetrate the cell directly, and the second occurs with the participation of mediators like tumor necrosis factor (TNF) 1. Heat shock, stress, free radicals, ultraviolet radiation, drugs, synthetic peptides, toxins, and numerous enzymes are examples of agents in the first process 6. Zhang et al. demonstrated for the first time that apoptosis was induced by TNF-α in an in vitro steatotic hepatocyte model 1.
Cells are complex genetic machines that once activated directly or indirectly enter into an apoptotic cycle. The cycle begins with the activation of a caspase -a cysteine protease that mediates intracellular proteolytic cascade and cleaves the cellular substrate and activates transcription factors 7. Once active, the transcription factors engender the expression of specific genes that determine the production of pro-apoptotic and anti-apoptotic factors 5.
In liver, excess fatty acid mobilization or lipid transport leads to apoptosis. Palmitate, a saturated fatty acid, has well-studied lipotoxic effects. The inability to convert fatty acid to triglycerides raises the risk of hepatocyte apoptosis and can probably be avoided by the use of an antioxidant. Hepatocytes are injured because the liver receives more fatty acids than can be oxidized or transported by VLDL 8-11. Hepatocyte damage begins with modification in mitochondrial morphology from fission to fusion, which decreases cellular adenosine triphosphate (ATP) levels and increases reactive oxygen species (ROS) 12.
Oxidative stress results from an imbalance between substances that promote oxidation and others with antioxidant effects; the consequence is a cellular lesion when pro-oxidants predominate 13. ROS are pro-oxidant substances that are responsible for intracellular lipid peroxidation that produces malondialdehyde (MDA) and 4-hydroxy-2-nonenal (HNE). It alters the protein, lipid, polysaccharide, and DNA synthesis 14,15.
The objective of this study is to discuss if and how modifications in a metabolic pathway due to a high-protein, high-fat, and low-carbohydrate diet can increase liver damage.
METHODS
ANIMALS
The study received prior approval by the Institutional Review Board for Animal Research (CEUA), Fluminense Federal University, case number 648, February 27, 2015. The study was conducted in the Experimental Nutrition Laboratory of the School of Nutrition, Fluminense Federal University (UFF), Niterói, RJ, Brazil. Animals were obtained from the Laboratory Animal Facility of the Oswaldo Cruz Foundation, Ministry of Health, Rio de Janeiro, Brazil.
After five days of adaptation, the rats were separated into two groups: the control diet group (CDG) (n = 9) and the experimental diet group (EDG) (n = 8). They were housed in individual polypropylene cages with controlled temperature (24 ± 2 oC) and humidity (60 ± 10%) and an alternating light-dark cycle (06:00 AM to 18:00 PM and 18:00 PM to 06:00 AM).
DIETS
Pragsoluções Biociências Comércio e Serviços, LTD, Jaú, São Paulo, Brazil handled the preparation of the diets. The control diet (AIN-93-M) had the following composition: carbohydrate (76.98), protein (13.56), and fat (9.46%). The experimental diet was based on the Atkins formula, with carbohydrate (8.77%), protein (59.46%), and fat (31.77%). Tert-butylhydroquine was calculated as 0.002 mg/1 g fat. The diets had the same amounts of vitamins, minerals, L-cystine, choline, and fiber, based on AIN-93-M specifications 16. The animals received diet and water ad libitum for eight weeks.
EXPERIMENTAL PROCEDURES AND SAMPLE COLLECTION
The estrous cycle phase was investigated, and animals in estrus were separated and fasted for eight hours. The animals were anesthetized by intraperitoneal injection of a solution containing 11.5 mg/100 g body mass of ketamine and 0.1 mg/100 g body mass of xylazine 17.
A capillary blood sample was drawn from the tails and blood samples from cardiac puncture. Samples were separated into two tubes: one with sodium heparin and the other with 0.7% EDTA with 4-(2-aminoethyl) benzenesulfonyl fluoride hydrochloride (AEBSF) 1 mg/mL of blood from Sigma-Aldrich, USA. The blood was centrifuged for 20 minutes at 314 rad.s-1 and stored at -80 oC for analysis.
The livers were removed and washed with saline solution, and flow cytometry analysis was performed immediately using the FITC Annexin V Apoptosis Detection Kit I components: 10X Annexin V Binding Buffer; FITC Annexin V; Propidium Iodide Solution from BD Pharmingen(tm); and flow cytometer FACF-CALIBUR BD model.
Intra-abdominal adipose tissue was carefully removed and weighed with a BioPrecisa(r) precision scale.
ANALYTIC METHODS
Capillary blood samples were collected for glucose measurement using the handheld Johnson & Johnson OneTouch(r), Ultra Mini(tm) meter. Plasma triglycerides in heparinized tubes were measured using automatic analysis (Vitalab Selectra E) with commercial kits from BioSystems Reagents & Instruments in the Multidisciplinary Research Support Laboratory (LAMAP), School of Medicine, UFF, Niterói, RJ, Brazil. The β-hydroxybutyrate and MDA in heparinized tubes were measured using the Beta-Hydroxybutyrate (beta HB) Assay Kit (ab83390) and Lipid Peroxidation (MDA) Assay Kit (ab118970), respectively, from Abcam(r), USA. The plasma was filtered through 10 kDa Spin Filter (ab93349) from Abcam(r), USA, to remove interfering substances, and quantified colorimetrically for MDA λ = 532 nm and β-hydroxybutyrate λ = 450 nm with a SpectraMax Plus 384 Microplate Reader. The analyzes were done at the Multidisciplinary Research Support Laboratory, School of Medicine, UFF, Niterói, RJ, Brazil. Insulin, glucagon, IL-6, and TNF-α in EDTA tubes were measured using Multiplex Biomarker Immunoassays for Luminex xMAP technology (Millipore, Billerica, MA, USA) lot number 2634193. Measurements were performed three times. The analyses were performed at the Specialized Laboratory for Clinical Analyses, São Paulo, Brazil.
STATISTICAL ANALYSIS
Data are presented as means ± standard deviations. Differences between the groups were analyzed using the Student's t-test, and the values were considered unpaired and parametric. An F-test was used to compare variances. Correlations between MDA and triglycerides and nonviable liver cells were analyzed with Pearson's correlation coefficient. Statistical significance was set at p < 0.05. The software package was GraphPad-Prism version 6.0e for Mac OS X, 2015.
RESULTS
No statistically significant differences were found between the groups for capillary glucose, insulin, glucagon, IL-6, or TNF-α. The β-hydroxybutyrate and MDA were expressed at the highest levels (p = 0.001 and p = 0.009, respectively) and triglycerides at the lowest levels (p = 0.005), in the experimental group. The percentage of nonviable cells in the experimental diet group was 2.18 times that of the control group (p = 0.001) (Table I).

The adipose tissue of rats fed on the experimental diet weighted less than the adipose tissue of rats that received the control diet. Mean EDG weight was 6.926 ± 1.118 g and mean CDG weight was 12.10 ± 1.849 g (p = 0.0339).
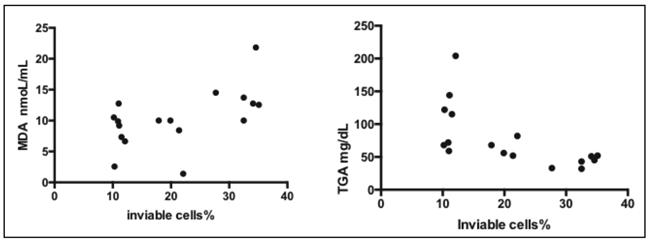
The results show a positive and significant correlation between the percentage of nonviable cells and MDA nmoL/mL levels (95% confidence interval, p = 0.0217) and a statistically significant negative correlation with triglycerides (TGA) mg/dL (p = 0.006) (Fig. 1).
DISCUSSION
Although the macronutrients composition of the experimental diet is not adequate for rats, it is carbohydrate content that regulates gluconeogenesis 18. Pichon et al. considered carbohydrate levels the most important factor in lipid metabolism 19 (Fig. 2).

Figure 2 The metabolic pathway responsible for hepatic damage associated with low-carbohydrate diet, high protein and high fat diet.
The results showed no statistically significant differences in glucose, insulin, or glucagon levels between the groups. The glucose levels are preserved by gluconeogenesis 20.
Low insulin and high glucagon may not occurs when gluconeogenesis is associated with high protein amounts that increases the secretion of insulin and glucagon 21-23. Our results are similar to the datas observed by Blouet et al. 23. Bielohuby et al. found low fasting glucose and low insulin levels associated with low carbohydrate and protein, and high fat diet 24. Westman et al. reported lower glucose, insulin, and C-peptide in humans receiving high fat versis high carbohydrate diet 20.
Corroborating previous studies, the adipose tissue weight was lower in the experimental diet than in the control 19,21,23,25,26. Low-carbohydrate decreased fat reserves and high-protein increased energy expenditure as protein needs more energy for digestion and metabolism 26,27. In adipose tissue, the lipolysis of triglyceride occurs with the supply of glycerol that will be converted to glucose, and fatty acids 28. In the liver, the fatty acids enter in the mitochondria, but are not stored as triglycerides, in the cytoplasm, because lipogenesis is inhibited 10,21,29. Botion et al. showed that rats fed with high-protein and carbohydrate free diet had a reduction of fat acids synthesis and low triglycerides in liver and plasma 30. Monteiro et al. demonstrated that hepatic steatosis was not found in rat livers that received high-fat, high-protein and low-carbohydrate diet. The diet composition was the same as the present work 31.
The plasma triglyceride levels in the present work are lower in the experimental group than the controls (Fig. 2). Kennedy et al. reported decreased plasma and hepatic triglycerides associated with a low-carbohydrate diet in rats fed on a ketogenic diet 32. Westman et al. conducted a thematic review with humans and found similar results 20.
In the mitochondria, fatty acids from the adipose tissue and the high-fat diet are broken down to produce energy through -oxidation. When the concentration of acetyl-CoA is elevated, it is converted to ketoacids that can be used as fuel for the brain, muscle, and heart 33. The pyruvate formed by an excess supply of amino acids from a high-protein diet enters the hepatocytes by a carrier-mediated transport. In the mitochondria, the pyruvate is converted to oxalacetate that may take two metabolic pathways -one to glucose be formed (the main one) or another to acetyl-CoA. When the offer of amino acids is elevated the second pathway becomes the mainly one 34. These data confirm this: the β-hydroxybutyrate level was 2.7 times higher in the experimental diet group than controls. This result supports the conclusion that gluconeogenesis is the main metabolic process and that low-carbohydrate diet is a ketogenic diet.
Kim et al. reported that when β-oxidation is abnormal (uncontrolled), levels of intermediate products increase and lead to inadequate energy production with a decrease in the NAD/NADH ratio and ATP. Consequently, lipid peroxidation occurs. This biochemical cascade leads to cellular membrane damage via the formation of reactive oxygen species (ROS), causing mitochondrial damage, oxidative stress and apoptosis 35. The elevated levels of β-hydroxybutyrate are another cause of lipid peroxidation and its consequences 36.
In our research the plasma MDA levels were higher in experimental diet versus the AIN-93M diet demonstrating the presence of lipid peroxidation (Fig. 2). The current study showed a positive correlation between the percentage of nonviable liver cells (apoptosis + non-apoptotic death) and MDA levels and a negative correlation with TGA levels (Fig. 1). This result suggests plasma MDA and triglycerides as probable indicators of hepatocyte damage.
Malondialdehyde (MDA) may reach distant tissues and extend cellular damage causing an increase in pro-inflammatory cytokines such as IL-6 and TNF-α 14. Here, plasma IL-6 and TNF-α levels did not differ significantly between the two groups. Oarada et al. found similar results when they fed rats on a low-carbohydrate, high-protein diet 37. Ellenbroek et al. showed that rats receiving a low-carbohydrate diet for 12 weeks had increased plasma IL-6. The diet in this study consisted of low protein and high fat (93.1%) 38.
In conclusion the metabolic pathway leads to modifications on plasma MDA and triglycerides doing them indicators of liver damage in rats fed with an experimental Atkins-type diet containing 59.46% of protein, 31.77% of fat and 8.77% of carbohydrate.

