










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkArchivos de Zootecnia
versión On-line ISSN 1885-4494versión impresa ISSN 0004-0592
Arch. zootec. vol.58 no.223 Córdoba sep. 2009
Análise multivariada da microflora epífita em S. Officinarum ensilada com G. Sepium*
Multivariate analysis of the epiphytic microflora on Saccharum Officinarum silages added with Gliricidia Sepium
Cabral Jr., C.1A, D. Silva2A, E. Amorim1B, W. Esfrain2B, E. Miranda1C, D. Pinheiro1D e J. Carnauba3
1Universidade Federal de Alagoas, UFAL. Maceió, Alagoas. CEP 57051-540. Brasil. Acyrorcjr@gmail.com; Bepra@fapeal.br; Cedmacdm@gmail.com; Ddmpinheiro@uol.com.br
2Universidade Federal da Paraíba-UFPB. Areia, Paraíba. CEP 58397-000. Brasil. Adivan@cca.ufpb.br; Bwep@cca.ufpb.br
3Universidade Federal Rural de Pernambuco-UFRPE. CEP 50050-540. Brasil. jcarnauba@hotmail.com
*Apoio FAPEAL. Parte da tese de doutorado do primeiro autor.
RESUMO
Objetivou-se com este trabalho caracterizar a microflora epífita da cana-de-açúcar (C), da gliricídia fresca (GNE) e emurchecida (GE) e das misturas ensiladas. Este estudo foi desenvolvido na Universidade Federal de Alagoas, UFAL, de janeiro a dezembro de 2005. O delineamento experimental foi o inteiramente casualizado em esquema fatorial [(2x3x4)+4]. As silagens de cana-de-açúcar aditivadas com GNE e GE foram confeccionadas nas proporções: 100/0, 75/25, 50/50 e 25/75. Os 28 tratamentos, com três repetições cada, foram acondicionados em silos experimentais (baldes plásticos de 10 l), vedados com lona de PVC e mantidos sob condições controladas de temperatura, umidade e proteção da presença de roedores. Os tempos de armaze-namento das silagens foram: 15, 45, 90 e 120 dias, nos quais foram abertos para a coleta de amostras. A análise multivariada mostrou-se significativa para os efeitos dos fatores e variáveis analisadas (p<0,1). A avaliação microbiológica das silagens baseou-se nos dois primeiros componentes principais (Yi), explicando 83,09% da variação total. As leveduras, os fungos e as bactérias totais, incluídos no Y1, explicaram 64,23%. Os bacilos tiveram menor importância para explicar a variabilidade na microflora epífita das silagens, ficando no Y2 e explicaram somente 18,86% da variação total. A análise de otimização forneceu as quatro melhores misturas em relação aos fatores e variáveis analisadas. Desta forma, 25% de gliricídia emurchecida podem vir a ser um valor controlador da fermentação microbiana indesejável em silagens de cana-de-açúcar.
Palavras chave: Bacilos. Bactérias. Fungos. Leveduras.
SUMMARY
The objective of this work was to evaluated the epiphytic micro flora of sugarcane (Saccharrum officinarum L.), fresh gliricidia (Gliricidia sepium) (GNE), wilted gliricidia (GE) and mixtures ensiled. This study was conducted at the Universidade Federal de Alagoas-UFAL, from January until December 2005. The experimental design was entirely randomized in outline [(2x3x4)+4)]. The sugarcane silages added with GNE or GE were maded in the referred percentages: 100/0; 75/25; 50/50 and 25/75. There were 28 treatments, each one with three replications. Silages were kept in experimental silos (plastic boxes), sealed with sailcloth and adhesive ribbon. These silos were maintained under controlled temperature, and humidity conditions and saved from rodent presence. The silos were opened at 15, 45, 90 and 120 days, and sampled. In relation to effects of factors and variables tested, the multivariate analysis showed significant differences (p<0.1). The microbiologic evaluation of silages was based in the two first principal components (Yi), which explained 83.09% of the total variation. The yeast, fungus and total bacteria, included in the Y1, explained 64.23%. The bacilli had less importance to explain the epiphytic micro flora variability of the silages, and were included in the Y2, explaining only 18.86% of the total variation. The 25% of wilted gliricidia may be an effective additive and it could be used with success to control undesirable secondary fermentation in sugarcane silages.
Key words: Bacilli. Bacteria. Fungus. Yeasts.
Introdução
Segundo Pedroso (2003), os microrga-nismos naturalmente presentes nas plantas forrageiras, chamados de microflora epífita, são responsáveis pela fermentação das silagens, afetando também a sua estabilidade aeróbia e a eficiência dos inoculantes microbianos. O número de microrganismos epífitos é variável, de acordo com o tipo de forragem, estádio de maturidade das plantas, clima, corte e condicionamento das forrageiras (Lin et al., 1992), bem como pela ocorrência de incêndio prévio (Bernardes et al., 2002; McDonald, 1991).
Geralmente, os microrganismos existentes em maior número nas plantas forrageiras são as enterobactérias, as leveduras e os fungos, que competem com os lactobacilos pelos açúcares durante a ensilagem, sendo considerados indesejáveis (Bolsen et al., 1992; Hendersom, 1991; Woolford, 1990). Além disso, a presença de leveduras, na ordem de 106 UFC/g de forragem (Alli et al., 1983) é prejudicial ao processo de ensilagem, porque estes microrganismos não contri-buem para a acidificação e estão associados com a deterioração aeróbia das silagens (Driehuis e Wikselaar, 1999) e não são inibidas pelo pH normalmente encontrado nas silagens. A população de bactérias láticas epífitas é muito variável; oscila de nenhuma a diversos milhões de unidades formadoras de colônias (UFC) por grama de forragem (Satter et al., 1988). Tem-se estimado que o número e tipos apropriados de organismos que se encontram na natureza são menores que 25% do ideal que se pode obter (Pedroso, 2003).
Ainda são poucos os trabalhos que ava-liam o efeito de aditivos vegetais ricos em proteína e carboidratos nas silagens de cana-de-açúcar. Também são escassos os estudos sobre a contagem de leveduras nestas mesmas silagens, podendo ser citados os de González e McLeod (1976) e Alli et al. (1983) e raríssimos os que procuraram caracterizar a população epífita da cana. Dentre estes, pode-se citar o de Lopez et al. (1998).
É necessário considerar que além das alterações na composição química da silagem, o desenvolvimento de fungos (Hendersom, 1991; Woolford, 1990) pode ser prejudicial à saúde dos animais e das pessoas. Segundo Edds (1983), todos os animais são sensíveis à ação das micotoxinas (Scudamore e Livesey, 1998; Amigot et al., 2006), podendo ocorrer lesões agudas que são provocadas pela ingestão de doses relativamente elevadas destas, ocasionando lesões hepáticas graves de efeito quase sempre letal, e, ou, crônicas, que produzem lesão progressiva, severa depressão no crescimento corporal e alterações hepáto-mas após período prolongado de ingestão destes volumosos. Dentre estas doenças, a aspergilose - causada por toxinas produzidas por Aspergillus flavus -, é bastante conhecida pela comunidade científica e rural. Segundo Sabino (1987), os gêneros Aspergillus, Penicillium e Fusarium, capazes de crescer em diversos substratos e sob condições ambientais variáveis, são produtores de micotoxinas (Scudamore e Livesey, 1998; Amigot et al., 2006) que ocasionam patologias nos animais. Os fungos Aspergillus glaucus e Aspergillus fumigatus, ambos patogênicos, produzem esporos que provocam doença respiratória maléfica em eqüinos e aves, além de causar problemas digestivos com o comprome-timento do desempenho destes animais (Pregnolato, 1985).
Já em relação aos organismos do reino Monera, Muck (1992) cita que os gêneros Clostridium e Listeria, como responsáveis por graves problemas em animais, podendo ocasionar diminuição no consumo voluntário, perda de desempenho e ou levá-los à morte. Dentre as patologias mais conhe-cidas, López et al. (1998) citam o botulismo e a listeriose.
Quanto à presença e desenvolvimento das leveduras, algumas são preocupantes em relação à saúde animal e humana, podendo ainda elevar as perdas de matéria seca da silagem pela produção de etanol. Dentre elas, podem ser citadas as dos gêneros ácido-metabolizantes como Candida e Hansenulla - as mais preocupantes -, e as fermentativas Torulopsis e Saccharomyces (Mahanna, 1994).
A gliricídia [Gliricidia sepium (Jacq.) Kunth ex Walp.] é uma leguminosa originária da América Central e já é cultivada em várias regiões do Brasil. Esta planta, além de possuir alto valor protéico (Atta-Krah, 1987), sintetiza fitoquímicos que atuam como poderosos agentes micostáticos, bactericidas e malaricidas (Ignatushchenko et al., 1997), o que sugere ter um grande potencial no controle de fermentações indesejáveis em silagens de cana-de-açúcar.
Com base no exposto, objetivou-se com este trabalho caracterizar a microflora epífita das silagens de cana-de-açúcar adicionadas com gliricídia.
Material e métodos
Este experimento foi conduzido no Laboratório de Fitopatologia e Fitossa-nidade, da Universidade Federal de Ala-goas-UFAL, de janeiro a dezembro de 2005. A cana-de-açúcar var. RB-92579 foi cedida pelo Programa de Melhoramento Genético da Cana-de-açúcar-PMGCA/UFAL. Os ramos jovens de gliricídia foram colhidos numa área já existente da Universidade Federal de Alagoas-UFAL. A cana-de-açúcar des-folhada e sem palhada não sofreu présecagem, ao passo que os ramos jovens da gliricídia colhidos foram utilizados sob o estádio fresco ou emurchecido, por aproximadamente 6 horas de exposição ao sol. Foram coletadas amostras dos componentes da mistura antes da ensilagem para posteriores análises.
O delineamento experimental foi o inteiramente casualizado em esquema fatorial [(2 x 3 x 4) + 4] em três repetições, sendo o fator A, relativo à gliricídia in natura (GNE) ou emurchecida (GE), adicionada à cana-de-açúcar (C) com percentuais iguais a 25, 50 e 75% (fator B); períodos de armazenamento do material ensilado iguais a 15, 45, 90 e 120 dias (fator C); além de mais quatro silagens de cana-de-açúcar sem aditivo (fator D), também em triplicata.
A mistura ocorreu após trituração em máquina forrageira com tamanho de partícula de aproximadamente 1,0 cm para cana-de-açúcar e de 2,5 cm para a GNE ou GE. Procurou-se manter uma densidade média de 450 kg/m3 durante o processo de compactação do material. Os silos foram identificados, vedados utilizando-se lona plástica e fita adesiva e armazenados em galpão sob temperatura e umidade controladas, além da proteção contra roedores.
Durante a abertura dos silos foram colhidas amostras (±25 g) de cada uma das 84 silagens, acondicionadas em tubos de ensaio previamente esterilizados, fechados e mantidos resfriados em caixas de polies-tireno (isopor) contendo bolsas de gelo, para serem encaminhados para o laboratório.
O isolamento e a contagem dos micror-ganismos deram-se segundo Lin et al. (1992). Amostras das silagens (25,0 g) foram pesadas em béqueres esterilizados contendo 250 ml de água destilada, trituradas em liquidificador e filtradas em papel de filtro Whatman No 10. As diluições em série (partindo-se de 10-1 até 10-4) foram realizadas adicionando-se 1 ml do extrato filtrado obtido em tubos de ensaio contendo 9 ml da solução de cloreto de sódio (NaCl) a 0,85%. Em seguida as amostras foram homogeneizadas e a inoculação foi realizada em placas de Petri contendo os meios de cultura seletivos para cada um dos microrganismos. Para quantificação de bacilos, adotou-se a mesma metodologia (Lin et al., 1992), no entanto, para a inoculação destes, retirou-se os tubos de ensaio do aparelho de ultra-som após 20 minutos colocando-os em banho-maria a 80oC.
Após a obtenção dos dados, aplicou-se a análise de componentes principais e de agrupamentos por Tocher. Para a escolha das silagens com os valores ótimos, a otimização foi baseada no índice de desejabilidade (experimentos com misturas), sugerido por Derringer e Suich (1980). Para a obtenção dos valores ótimos de cada variável analisada neste experimento, adotou-se intervalos sugeridos por Ma-hanna (1994). Todas as variáveis analisadas não apresentaram distribuição normal e ou homocedasticidade. Sendo assim, foram realizadas as transformações:
Inversa [y´= 1 / (y + k)] com λ igual a -1,0 e k= 74032,4 (leveduras); raiz quadrada [√X + 0,6] com λ igual a 0,5 e k= 0,6 (fungos); raiz quadrada [√X + 0,5] com λ igual a 0,5 e k= 0 (bacilos e bactérias totais).
Resultados e discussão
Os dados obtidos quanto à análise microbiana da cana-de-açúcar, gliricídia fresca e nos emurchecidos por 6 horas antes da ensilagem encontram-se apresentados na tabela I.

A tabela II apresenta através as equações de regressão e os respectivos coeficientes de determinação para leveduras, fungos, bacilos e bactérias totais. A apresentação dos resultados foi realizada considerando-se separadamente o aditivo fresco (GNE) e o que sofreu emurchecimento (GE). O teste usado para a significância dos coeficientes de regressão foi o F de Snedecor (p<0,1). Para manter a hierarquia dos termos de algumas das equações de regressão, alguns destes foram adicionados às referidas equações, mesmo apresentando-se não-significativos.
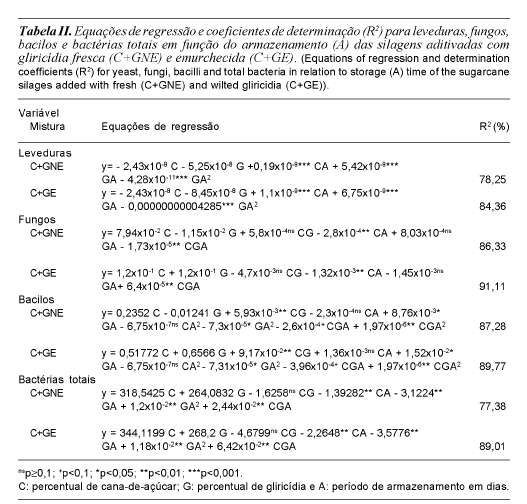
Observa-se que, em relação ao emurche-cimento do aditivo vegetal utilizado nas silagens de cana-de-açúcar, houve efeito significativo (p<0,05), visto que a análise baseada na técnica multivariada experimentos com misturas gerou modelos preditivos tanto para as que receberam gliricídia fresca (GNE) quanto para as que receberam a pré-seca (GE) ao sol por 6 horas aproximadamente (tabela II).
Em relação às leveduras, as silagens de C + GNE apresentaram diferença significativa para a interação cana (C) e armazenamento (A), com incremento positivo no número de unidades formadoras de colônia por grama de matéria verde (UFC/g MV) na ordem de 2,94 x 10-4 ponto percentual a cada dia de armazenamento e incremento percentual de cana. Ao passo que, com relação à interação gliricídia fresca (GNE) e armazenamento, ocorreu o inverso, ou seja, um decréscimo na população de leveduras na ordem de 2,56 x 10-5 ponto percentual. Estes resultados indicam que há influência da adição da gliricídia na silagem de cana-de-açúcar devido ao seu poder micostático através de seu metabolismo secundário.
Os resultados para a adição de GE nas silagens de C mostram que o desenvol-vimento da referida população de leveduras foi menor em relação à interação CxA (3,42 x 10-4%) e GxA (1,28 x 10-49%), indicando que ao aumentar o teor de matéria seca nas silagens, aumenta-se a pressão osmótica do meio e consequentemente, dificultou a proliferação destes microrganismos ao longo do armazenamento.
Para a população de fungos observa-se que à medida que aumentou o percentual da GNE nas silagens de cana-de-açúcar, houve diminuição significativa quadrática (p<0,05) na referida população destes microrga-nismos, onde foi observada ausência ou seus esporos estavam em latência.
Em relação ao armazenamento, observase que nas silagens exclusivas de cana-de-açúcar houve diminuição linear (p<0,05) da ordem de 2,80 x 10-4 ponto percentual a cada dia de armazenamento. Nas silagens com maior adição de GNE o percentual de fungos sofreu incremento linear (p<0,05) chegando a valores próximos de 4,5%. Aos 120 dias de armazenamento das silagens pode-se verificar menor porcentagem fúngica para a relação percentual 50:50, mas que aumentou a partir deste ponto, o que sugere que esta seja a melhor relação quando o aditivo não for pré-secado. Quando a adição foi feita com GE nas silagens de cana-de-açúcar, ocorreu o mesmo efeito negativo e quadráti-co para fungos (p<0,05), onde houve dimi-nuição de 1,32 x 10-3% para a interação CxA e nenhuma diferença (p<0,05) para a interação GxA.
Para a quantificação de bactérias totais observaram-se maiores valores (24145,0 UFC/g MV) para as silagens exclusivas de cana-de-açúcar e decréscimo linear (p<0,05) de 1,39% a cada dia de armazenamento. À medida que se aumentou o percentual de GNE nas silagens de cana-de-açúcar observou-se um decréscimo de 89,7% na população destes microrganismos até a proporção C/GNE atingir 25/75%, quando atingiu-se um valor de 8009,0 UFC/g MV. Quando o aditivo sofreu emurchecimento, houve o mesmo comportamento, porém a diminuição dos valores referentes à quantificação de bactérias totais foi de aproximadamente 25%. As interações CxA e GxA foram significativas e negativas (p<0,01) e influenciaram no número de UFC/ g MV das silagens na ordem de 2,26 e 3,57%, a cada adição de GE e dia de armazenamento, respectivamente. Mais uma vez, obteve-se maior população de bactérias totais nas silagens exclusivas de cana-de-açúcar, sugerindo que o aditivo vegetal utilizado também pode controlar estes microrganismos.
Para a população de bacilos nas silagens exclusivas de cana-de-açúcar não se observou variação no número de unidades formadoras de colônias por grama de matéria verde. Para as que receberam GNE e GE houve decréscimo à medida que se aumen-tou o percentual do referido aditivo e o armazenamento das respectivas silagens. Ficou evidenciado que a gliricídia pode ser utilizada como controladora de bactérias degradadoras de proteínas, as chamadas comumente de proteolíticas, ao observar os valores encontrados para os coeficientes referentes às interações CA2, GA2 e CGA, -6,75 x 10-7, -7,30 x 10-5 e -2,59 x 10-4, respectivamente, para ambos os teores de MS da gliricídia.
Nas silagens com adição de 75% de GNE se observou as menores populações de bactérias totais tanto nos primeiros dias quanto nos últimos dias após a ensilagem do material. Quando a gliricídia sofreu emurchecimento, os resultados sugerem que houve contaminação do material ao entrar em contato com o solo. O incremento foi positivo e linear (p<0,01) e se deu em maior destaque aos 120 dias após a ensilagem. Neste caso, os resultados sugerem que, mesmo com o aumento da pressão osmótica, do percentual de GE nas silagens e do tempo de armazenamento, a população de bacilos não foi controlada pelos metabólitos secundários da gliricídia e que provavel-mente houve perdas de matéria seca e valor nutritivo do material ensilado.
Percebe-se que a gliricídia influenciou significativamente na microflora epífita das silagens analisadas neste estudo, já que a mesma sintetiza fitoquímicos (fenóis, saponinas, xantonas, dentre outros) que podem atuar como poderosos agentes fungicidas e bactericidas (Ignatushchenko et al., 1997).
A tabela III apresenta os coeficientes de correlação de Pearson (r) para as variáveis analisadas durante a armazenagem das 28 silagens de cana-de-açúcar aditivadas com gliricídia fresca ou emurchecida por seis horas ao sol.

Observou-se correlação linear e direta-mente proporcional para leveduras e fungos (r= 0,52; p<0,01) e leveduras e bactérias totais (r= 0,67; p<0,01). Em relação a fungos e bacilos, observou-se correlação linear inversamente proporcional (r = -0,49; p<0,01), o que sugere algum tipo de controle populacional por parte dos fungos estabe-lecidos nas silagens analisadas sobre a população de bacilos. Para a correlação linear entre bactérias totais e bacilos observou-se a mesma tendência (r= -0,38; p<0,05). Isto sugere que a acidificação dos substratos causada por determinados gêneros incluídos no grupo das bactérias totais (hetero e ou homofermentativas) pode ter exercido efeito controlador sobre os bacilos.
SIMILARIDADE MICROBIOLÓGICA
A avaliação da similaridade microbiana das 28 silagens em estudo baseou-se nos dois primeiros componentes principais que explicaram 83,1% da variação total.
Com base no princípio de que a impor-tância relativa dos componentes principais (Jolliffe, 1986) decresce do primeiro para o último, tem-se que os últimos componentes são responsáveis pela explicação de uma fração mínima da variância total disponível (Cruz e Regazzi, 1997). É possível verificar na tabela IV que os bacilos (0,98) foram a variável que apresentou maior coeficiente de ponderação (elemento do autovetor), em valor absoluto, nos componentes de menor autovalor (menor proporção de variação explicada), sendo assim, considerada de menor importância para explicar a varia-bilidade microbiológica das silagens de cana-de-açúcar relacionadas às fermen-tações primária (desejável) e secundária (indesejável).

A dispersão dos escores referentes à posição de cada tratamento (24 silagens de cana-de-açúcar aditivadas com GNE ou GE mais as quatro silagens sem aditivo), em relação a sua ortogonalidade cartesiana, pode ser observada após a classificação final, onde os tratamentos 7, 8, 10, 11, 12, 14, 18, 21, 25 e 26 e os tratamentos 4, 15, 16, 19, 20, 22, 23, 24, 27 e 28 compuseram, respectivamente, os grupos Y1 e Y2, devido à menor dispersão dos escores nos dois primeiros componentes principais, tendo sido considerados dentro de cada grupo, como os mais similares.
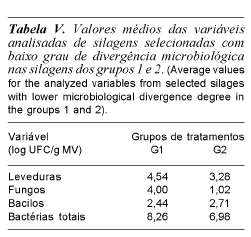
A tabela V apresenta os valores médios, relativos às variáveis leveduras, fungos, bacilos e bactérias totais para os dois primeiros grupos de tratamentos. A mesma explica -em termos de média aritmética-, que praticamente a população de bacilos manteve-se a mesma nos dois primeiros grupos, ou seja, os respectivos tratamentos apresentaram populações de bacilos semelhantes corroborando a hipótese de que foram controlados pelos outros microrganismos durante a fase anaeróbica.
OTIMIZAÇÃO EXPERIMENTAL
De acordo com Bruns et al. (2007) é através de um planejamento experimental que se obtém com custo e tempo mínimos, as informações que se desejam a respeito do efeito das proporções de cada componente presente na mistura sobre as características do produto final resultante desta mistura. Além disto, conhece-se o erro experimental associado à informação de que se dispõe, podendo-se estabelecer o grau de confiança da mesma.
Heinsman e Montgomery (1995), tratando da otimização de produtos usando experimentos com misturas (EM) argumentam sobre a relevância de tais projetos. Estes autores ao citarem que o desenvolvimento de fórmulas para diversos produtos é tradicionalmente feito por tentativas e erros, tendo-se que sempre variar as proporções de um dos ingredientes por vez. Isto pode consumir muito tempo e não permite uma compreensão das interações que possam existir entre os diversos ingredientes.
Com base nesta técnica, definiu-se as quatro melhores misturas (tabela VI), permitindo assim, a continuidade das análises com maior economia e menor tempo. Os intervalos definidos para o ponto ótimo para cada uma das variáveis analisadas pelo pacote estatístico utilizado foram retirados de diversos trabalhos constantes na literatura científica.

A tabela VI mostra as quatro silagens que apresentaram valores ótimos dentro dos intervalos fixados a priori (para os componentes cana e gliricídia); e a posteriori (para os fatores emurchecimento e armazenamento; e variáveis analisadas) de acordo com o índice de desejabilidade (Derringer e Suich, 1980). Pode-se observar que houve predominância média de 75% de cana-de-açúcar + 25% de gliricídia.
O aditivo vegetal pré-secado por seis horas ao sol, apresentou-se como sendo a melhor alternativa para a confecção das respectivas silagens, porém, observou-se a presença de bacilos no material emur-checido. Este tipo de contaminação acontece normalmente quando o substrato utilizado para a pré-secagem da forragem contém uma quantidade de matéria orgânica rica em húmus ou serrapilheira, ou que foram anteriormente utilizadas para pastejo.
De um intervalo pré-fixado de 15 a 120 dias de armazenamento, houve predomi-nância média para as quatro primeiras silagens com períodos próximos de 110 dias. Respectivamente, as mesmas misturas (silagens) apresentaram índices de deseja-bilidade iguais a 1,0.
Cabral Jr. et al. (2007) citam que 108 UFC/ g MV de bactérias totais pode ser considerado um valor ideal para tal otimização. Segundo estes autores, tal sugestão baseou-se no fato de que, na maioria dos trabalhos envolvendo adição de inoculantes microbianos em silagens de cana-de-açúcar, so-mente em relação ao número de bactérias produtoras de ácido lático e ou ácido acético -excluindo-se outras espécies homo e heterofermentativas-, a quantidade normalmente encontrada é de 106 UFC/g MV, o que sugere que não está sendo considerada a microflora original das forragens envolvidas na ensilagem nem a provável contaminação por máquinas colheitadeiras e picadoras, e ou o substrato no qual as plantas são emur-checidas.
Desta forma, a técnica de otimização de misturas foi aplicada com sucesso, e análises posteriores, consideradas de maior impor-tância e de custos mais elevados -como as referentes à identificação das espécies epífitas mais patogênicas (Clostridium botulinum, Bacillus sp. Listeria monocy-togenes, Aspergillus flavus, Fusarium moniliforme, Penicillum ssp. Hansenula ssp. e Candida ssp.), das de maior benefício (Lactobacillus plantarum e Lactobacillus buchneri) e dos bioensaios utilizando-se extratos em diferentes níveis do aditivo vegetal (gliricídia) no controle destes microrganismos (por cromatografia de ca-mada delgada - TLC), sem deixar de ser considerado o fator tempo-, poderão ser realizadas com maior eficiência e acurácia nas quatros misturas/silagens indicadas.
Conclusão
A análise de experimentos com misturas através do indice de desejabilidade indicou que as melhores silagens de cana com diferentes níveis de gliricídia são as confeccionadas na proporção aproximada de 75 e 25% de cana e gliricídia emurchecida, com armazenamento em torno de 90 a 120 dias.
Bibliografia
Alli, I., R. Fairbairn and B.E. Baker 1983. The effects of ammonia on the fermentation of chopped sugarcane. Anim. Feed Sci. Tech., 9: 291-299. [ Links ]
Amigot, S.L., C.L. Fulgueira, H. Bottai and J.C. Basilico. 2006. New parameters to evaluate forage quality. Postharvest Biol. Tec., 41: 215-224. [ Links ]
Atta-Krah, A.N. 1987. Flowering and seed production of Gliricidia sepium. In: Workshop on Gliricidia sepium: Management and Improvement, 1997, Turrialba. Proceedings... CATIE. Turrialba; Nitrogen Fixing Tree Association. Hawaii. NFTA Special Publication, 87.01. p. 142-145. [ Links ]
Bruns, R.E., I.S. Scarminio e B. Barros Neto. 2007. l. Como fazer experimentos: pesquisa e desenvolvimento na ciência e na indústria. Editora da Unicamp. Campinas, SP. 480 p. [ Links ]
Bernardes, T.F., R.N. Silveira, R.M. Coan e R.A. Reis. 2002. Características fermentativas e presença de levedura na cana-de-açúcar crua ou queimada ensilada com aditivo. Em: 39 Reunião Anual da Sociedade Brasileira de Zootecnia. Recife, 2002. Anais... [ Links ]
Bolsen, K.K., C. Lin, B.E. Brent, A.M. Feyerherm, J.E. Urban and W.R. Aimutis. 1992. Effects of silage additives on the microbial succession and fermentation process of alfalfa and corn silages. J. Dairy Sci., 75: 3066-3083. [ Links ]
Cabral Jr., C.R., E.C. Miranda, D.M. Pinheiro, I.G. Gomes, M.V.M. Andrade e M.S.C. Pinto. 2007. Dinâmica fermentativa de silagens de Gliricidia sepium. Arch. Zootec., 56: 249-252. [ Links ]
Cruz, C.D. e A.J. Regazzi. 1997. Modelos biométricos aplicados ao melhoramento genético. 2a ed. Universidade Federal de Viçosa. Viçosa, MG. 390 p. [ Links ]
Derringer, G. and R. Suich, 1980. Simultaneous optimization of several response variables. J. Qual. Technol., 12: 214-219. [ Links ]
Driehuis, F. and P.G. Wikselaar. 1999. The occurrence and prevention of ethanol fermentation in high-dry matter grass silage. J. Sci. Food Agric., 80: 711-718. [ Links ]
Edds, G.T. 1983. Acute aflatoxicosis: Review. J. Amer. Vet. Med. Ass., 162: 304-309. [ Links ]
González, E. and N.A. McLeod. 1976. Spontaneous fermentation of sugarcane. Trop. Anim. Prod., 1: 80-84. [ Links ]
Heinsman, J.A. and D.C. Montgomery. 1995. Optimization of a household product formulation using a mixture experiment. Qual. Eng., 7: 583-600. [ Links ]
Hendersom, A.R. 1991. Biochemistry in forage conservation. Conference of Conservation Towards 2000. Brandschweig (23-25 Jun). [ Links ]
Ignatushchenko, M.V., R.W. Winter, H.P. Bachinger, D.J. Hinrichds and M.K. Riscoe. 1997. Xanthones as antimalarial agents: studies of a possible mode of action. FEBS Lett., 409: 67-73. [ Links ]
Jolliffe, I.T. 1986. Principal component analysis. Springer-Verlag. NewYork. 271 p. [ Links ]
Lin, C., K.K. Bolsen and R.A. Hart. 1992. Epiphytic micro flora on alfalfa and whole-plant corn. J. Dairy Sci., 75: 2484-2493. [ Links ]
López, Z.A., I.E. Moreño, F.A. Fogliata, H.G. Ayala and C. Tornec. 1998. Microbial population of sugar juice that is neither affected nor deteriorated by frost. Sugar y Azúcar, 83: 21-34. [ Links ]
Mahanna, W.C. 1994. Genetic selection for forage nutritional quality. In: Quality forage and ruminants; Proceedings... Ministry of Agriculture and Food/Guelph & Brockville. Ontario. [ Links ]
McDonald, P., A.R. Henderson and S.J.E. Heron. 1991. The biochemistry of the silage. J. Wiley and Sons. Edinburgh. 226 p. [ Links ]
Muck, E.C. 1992. The role of silage additives in making high quality silage. In: National Silage Production Conference. NRAES-67. Ithaca, N.Y., 1992. Proceedings. NRAES. Ithaca, N.Y. p. 106-116. [ Links ]
Pedroso, A.F. 2003. Aditivos químicos e microbianos no controle de perdas e na qualidade de silagem de cana-de-açúcar (Saccharum officinarum L.). Tese (Doutorado). Esalq. Piracicaba-SP. 120 p. [ Links ]
Pregnolatto, W.R. 1985. Normas analíticas do Instituto Adolfo Lutz. 3a ed. Instituto Adolfo Lutz. São Paulo, v. 1. [ Links ]
Sabino, M. 1987. Micotoxinas, ocorrência natural em alimentos: aspectos gerais. Instituto Adolfo Lutz. São Paulo/SP. p. 40-48. [ Links ]
Satter, L.D., R. Muck, J.A. Woodford and B.A. Jones. 1988. Inoculant research: what has it shown us?. In: Forage Production and Use Symposium, 12. Wisconsin Dells. Proceedings. Wisconsin Forage Council. Wisconsin Dells. p. 108-119. [ Links ]
Scudamore, K.A. and C.T. Livesey. 1998. Occurrence and significance of micotoxins in forage crops and silages: a review. J. Sci. Food Agric., 77: 1-17. [ Links ]
Woolford, M.K. 1990. The detrimental effects of air on silage. J. Appl. Bacteriol., 68: 101-116. [ Links ]
Recibido: 2-9-07.
Aceptado: 14-5-08.

