










My SciELO
 Custom services
Custom servicesServices on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActas Urológicas Españolas
Print version ISSN 0210-4806
Actas Urol Esp vol.33 n.9 Oct. 2009
Hipoxia tumoral. Papel del factor inducible por hipoxia
Tumor hypoxia: the role of HIF
Avelino Fraga, Ricardo Ribeiro y Rui Medeiros
Instituto Português Oncologia, Oncologia Molecular, Porto, Portugal
Dirección para correspondencia
RESUMEN
Los tumores sólidos, por lo general, existen y progresan en un ambiente de hipoxia; así se observa que las células tumorales son resistentes a la apoptosis y se acompañan de un aumento de la angiogénesis, volviéndose más agresivas, con capacidad invasora y resistentes al tratamiento.
La genética y los mecanismos biológicos subyacentes a este fenómeno son todavía poco claros, pero muchos estudios sugieren un papel del factor inducible por hipoxia (hipoxia inducible factor [HIF]) en este proceso. En condiciones de hipoxia, la subunidad alfa no es destruida y activará la transcripción de un conjunto de genes que contribuyen a la agresividad del tumor. Su expresión está asociada a un aumento del potencial metastásico que se verifica tanto en estudios animales, como en tumores humanos.
La hipoxia tumoral se ha convertido en un factor clave en la progresión tumoral y se asocia a un mal pronóstico, sobre todo en tumores de riñón y próstata. Este trabajo tiene por objetivo revisar la importancia de la hipoxia en la carcinogénesis y en la progresión tumoral, presentando una revisión de los conocimientos actuales sobre el tema, mecanismos de acción y la activación del HIF-1a.
Palabras clave: HIF, Hipoxia, Cáncer.
ABSTRACT
Solid tumors usually occur and progress in a hypoxic enviroment, suggesting that tumor cells are resistant to apoptosis and are associated to an increased angiogenesis, which makes them more aggressive, with invasive capacity and resistant to treatment.
The genetic and biological mechanisms underlying this phenomenon are still unclear, but many studies suggest a role of HIF in this process. Under hypoxic conditions, the alpha subunit is not destroyed, and will activate transcription of a set of genes contributing to tumor aggressiveness. Its expression is associated to an increased metastatic potential that has been shown in both animal studies and human tumors.
Tumor hypoxia has emerged as a key factor in tumor progression and is associated to a poor prognosis, particularly in kidney and prostate tumors. The purpose of this study was to review the significance of hypoxia in carcinogeneses and tumor progression by reviewing the current knowledge on the subject and the mechanisms of action and activation of HIF-1a.
Key words: HIF, Hypoxia, Cancer
Introducción
El factor inducible por hipoxia (hipoxia inducible factor [HIF]) es un factor de transcripción que regula la respuesta celular a la hipoxia y actúa como regulador de la homeostasis del oxígeno1-3. La identificación del sistema de transcripción HIF por Wang y Semenza4 es crucial para entender la fisiología del O2; sabemos actualmente que el HIF y la hipoxia son los mayores determinantes en la angiogénesis y que, por ejemplo, regulan los procesos de invasión y metastización determinantes de la agresividad tumoral.
El factor de transcripción activa genes que codifican las proteínas que aumentan la disponibilidad del oxígeno y permiten la adaptación metabólica en ausencia de oxígeno, controlando la expresión de decenas de productos de los genes y proteínas implicados en la angiogénesis, la eritropoyesis, la glucólisis, la invasión, la apoptosis, el tono vascular, la regulación del pH, la homeostasis epitelial y la resistencia a los fármacos.
Se identificaron más de 60 genes diana inducidos por HIF2, mientras que otros son suprimidos7; todavía muchas funciones son dependientes del HIF7.
Estructura molecular del HIF-1a
El gen HIF1A, que codifica el HIF-1a, se localiza en el locus 14q21-q249, que contiene 15 exones10. Es un heterodímero compuesto por cadenas alfa (reguladas por el O2) y beta, dispuestas en doble hélice (bHLH); pertenece a una familia de factores de transcripción que consta de tres subunidades alfa (HIF-1a, HIF-2a, HIF-3a) y a una subunidad beta (HIF 1b), también llamada translocador nuclear aril hidrocarburos ARNT)4,11,15.
Hay 2 locales de señalización nuclear (NLS), situados en el C-terminal (aminoácidos 718-721) y en el N-terminal (aminoácidos 17-33), pero sólo el C-terminal es el responsable por la acumulación nuclear del HIF-1a16. Además, se sabe que el HIF contiene 2 dominios de transactivación (TAD) en el C-terminal (aminoácidos 531-575 y 786-826), que están separados por una secuencia de aminoácidos (575-786) que inhiben la transactivación17 (fig. 1).

Figura 1 - Estructura molecular de HIF-1a. Adaptado de Shi YH55.
La parte N-terminal de la molécula (aminoácido 1-390) contiene el dominio bHLH-PAS, que es necesario para la dimerización y ligación al ADN18. Las interacciones entre los dominios bHLH de ambas subunidades regulan su dimerización19.
El dominio C-terminal tiene como función señalizar la translocación del HIF-1a para el núcleo, la estabilización proteica y la interacción con el coactivador p30017. En el dominio de la degradación oxígeno-dependiente (ODD) del HIF-1a, los residuos de prolina en las posiciones 402 y 564 tienen un importante impacto en la estabilidad de la proteína en condiciones de normoxia, pues permiten, cuando están hidroxiladas, el reconocimiento por la proteína Von Hippel Lindau (pVHL) y la posterior activación de la vía de degradación de la ubiquitina20-25. La hidroxilación de los residuos de prolina en el ODD del HIF-1a representa el punto crítico que regula la estabilidad de la proteína26,27 (fig. 2). La actividad de la transcripción de genes HIF1A se encuentra, de este modo, regulada por la tensión de oxígeno celular.

Figura 2 - Reglamento de la estabilidad y la actividad del factor inducible
por hipoxia. Adaptado de Brahimi-Horn y Pouyssegur94.
Mecanismos moleculares del HIF y de la activación del HIF1A
En presencia de O2, los dominios de hidroxilación de la prolina (PHD1, 2, 3) provocan la hidroxilación específica en dos residuos de prolina (P402 y P564) en el ODD del HIF-1a, permitiendo el reconocimiento del HIF-1a por la pVHL, formándose el complejo E3 ubiquitina, que transformará el HIF-1a en un objetivo para la degradación30-33. Jaakkola et al32 demostraron que la interacción entre la pVHL y el dominio específico del HIF-1a están regulados por la hidroxilación del residuo de prolina (HIF-1a P564) por una enzima denominada HIF-1a prolil hidroxilasa (HIF-PH), necesitando de hierro y oxígeno.
Otro sensor de O2 es el factor inhibidor del HIF-1 (FIH-1), que hidroxila el HIF-1a en presencia de O2, en el residuo de asparagina 803 en el dominio de la activación de la transcripción del C-terminal (C-TAD), y permanece inactivo en hipoxia, lo que permite la interacción con los coactivadores CBP/p30034,35 (fig. 2).
En condiciones de hipoxia, el O2 molecular no está disponible y de esta forma las enzimas están inactivas, lo que implica un aumento de los valores del HIF-1a36. El HIF-1a no es hidroxilado y, por tanto, degradado, provocando su acumulación en la forma heterodimizada con la subunidad beta (HIF-β). Este heterodímero emigra para el núcleo, donde se unirá a las secuencias específicas del ADN, activando genes implicados en la adaptación a la hipoxia, supervivencia celular, angiogénesis y metástasis, como, por ejemplo, el factor de crecimiento endotelial vascular (VEGF), el factor de crecimiento de transformación-alfa (TGF-a), el transportador 1 de glucosa (GLUT-1), o la anhidrasa carbónica IX (CA9), entre muchos otros, que sabemos están implicados en el desarrollo y la agresividad tumoral37,38.
Por tanto, el principal regulador del HIF es el oxígeno22,39. El segundo en importancia son los oncogenes, que pueden contribuir para estabilizar la proteína o degradarla. Por ejemplo, el producto del gen supresor tumoral TP53, proteína p53, inhibe la actividad del HIF-1a, volviéndose un objetivo para la degradación proteosómica40. Sin embargo, las deleciones o mutaciones de TP53 pueden facilitar la acumulación del HIF1a en situaciones de hipoxia, aumentando la expresión del VEGF por las células tumorales.
El producto del gen supresor tumoral VHL también regula la estabilidad del HIF-1a42, ya que en presencia de oxígeno la pVHL puede ligarse a la subunidad HIF-1α, transformándose en un objetivo para la prolil-hidroxilación25-27. Además, otros oncogenes (v-Src o RasV12) inhiben la prolil-hidroxilación, lo que implica estabilización del HIF-1a39-42.
También sabemos que la expresión del gen HIF1A puede regularse por otras vías, principalmente las vías de señalización intracelular, tales como la proteína cinasa B (Akt) y la 3- fosfatil-inositol-cinasa (PI3K), aunque todavía no está claro su papel en estas vías de regulación.
También están descritas otras moléculas reguladoras del HIF1A, tales como las especies reactivas de oxígeno (ROS) participantes en la carcinogénesis o citocinas, como el factor de necrosis tumoral (TNF-a) y la angiotensina49-53, bien como otras vías de señalización, como el RAS/RAF1/MEK1/ERK1/2 y/op53/JNK, que se activan en respuesta de los oncogenes, factores de crecimiento o hipoxia (fig. 3).
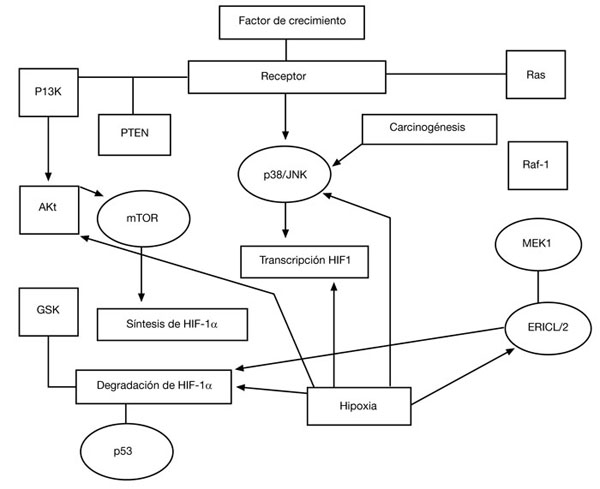
Figura 3 - Vías de señalización y regulación de HIF-1 α: oncogenes,
factores de crecimiento e hipoxia. Adaptado de Shi YH et al74.
Funciones generales del gen HIF1A
La hipoxia es la reducción de la tensión de oxígeno, que se define en términos clínicos por la reducción de la disponibilidad de oxígeno hasta niveles críticos, es decir, los valores de tensión inferiores al 7%53.
El HIF-1a está involucrado en la respuesta a la hipoxia y también en la homeostasis del oxígeno, en la respuesta a la isquemia del miocardio, isquemia cerebral, isquemia de la retina, hipertensión pulmonar, la preeclampsia, el retraso en el crecimiento intrauterino y el cáncer. Tiene un papel central tanto en los mecanismos fisiológicos homeostáticos, como en los etiopatológicos. Actúa sobre genes diana, dado que su función es regulada por factores de crecimiento y alteraciones genéticas implicadas en la progresión tumoral54,55.
Los vasos aberrantes pueden desaparecer en cualquier momento, pero a veces pueden reutilizarse, provocando reoxigenación local, estimulando cambios repentinos de hipoxia y reoxigenación como resultado de la angiogénesis local56-59.
El medio ambiente tumoral está bien caracterizado, si lo entendemos como una fluctuación de hipoxia y privación de nutrientes que lleva a la adaptación genética y epigenética de clones celulares, aumentado la capacidad de invasión y metastización.
Además, estas adaptaciones a la hipoxia vuelven a los tumores más difíciles de tratar y con mayor resistencia a las terapias. Una parte importante de este proceso consiste en la adaptación de los productos de los genes en respuesta a la hipoxia y muchos de estos genes reguladores de hipoxia son mediados por el HIF1A60; se estima que alrededor del 1% del genoma es regulado por la hipoxia.
La hipoxia tumoral por sí sola es un importante factor epigenético de la regulación de la proteína HIF-1a. Además de inhibir las PSD y al HIF-1a, la hipoxia genera radicales libres de oxígeno, que son capaces de estabilizar la proteína HIF-1a y de inducir los genes HIF y VEGF61,62.
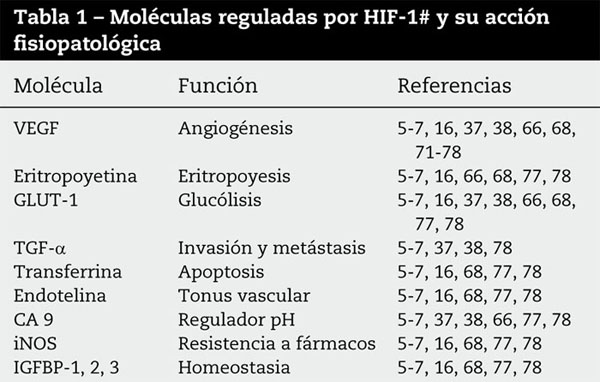
Cuando se establece la hipoxia, hay una respuesta celular para evitar la apoptosis63 y se activa el factor de transcripción HIF-1a, que genera un heterodímero con el HIF-1β(ARNT) en el elemento de la respuesta a la hipoxia (HRE), lo que lleva a una respuesta celular múltiple con activación de los oncogenes64, aumento de la vascularización con producción de VEGF, aumento del transporte de glucosa (GLUT1) y de la actividad de la anhidrasa carbónica (CA9) y aun a la inducción de varios genes apoptóticos65-67. Se sabe que el HIF actúa sobre los genes codificadores de la eritropoyetina, transferrina, endotelina-1, sintetasa inducible de óxido nítrico (iNOS), hemooxigenasa 1, factor de crecimiento insulínico-2 (IGF-2), proteína de unión 1, 2 y 3, al factor de crecimiento insulínico (IGFBP 1, 2, 3), transportadores de glucosa (GLUT) y enzimas glucolíticas18,28,68 (fig. 4), promoviendo la adaptación metabólica a la hipoxia, y es también regulado por la presión de O2, dependiendo de la expresión de la subunidad HIF-1a69. La capacidad de adaptación de la hipoxia por las células malignas es fundamental para el crecimiento tumoral (tabla 1).

Figura 4 - Respuestas determinadas por factor inducible por hipoxia: actúa
como el principal regulador fisiológico de la hipoxia. Adaptado de Acker y Plate93.
Hipoxia, factor inducible por hipoxia y cáncer
La hipoxia es significativamente menor en los tumores en que el promedio de la presión de O2 es superior al 1,5%53,79,80.
Las células tumorales, para sobrevivir, tienen que adaptarse a la baja presión de pO2; muchos productos genómicos están implicados en la neoangiogénesis tumoral. Estas adaptaciones contribuyen a la supervivencia fenotípica y la agresividad clínica81. La hipoxia tumoral se ha asociado a mal pronóstico en numerosos tipos de cáncer82.
Los clones de células tumorales tienen la capacidad de adaptarse a microambientes hipóxicos, ya sea en los locales primarios, como en los sitios de metástasis. Los mecanismos genéticos y epigenéticos de la adaptación a la hipoxia (inestabilidad genética, glucólisis aeróbica, la pérdida de control del ciclo celular, la pérdida de señales de la apoptosis) son característicos de malignidad60 (fig. 5).

Figura 5 - Reglamento de HIF-1a.
Hay pruebas de que la hipoxia puede controlar y mantener la inestabilidad genética. Esta inestabilidad genética puede disminuir la reparación del ADN y aumentar la tasa de mutaciones66.
La hipoxia intratumoral es un factor de mal pronóstico observado en tumores de próstata, mama, musculoesquelético, cabeza y cuello, y cérvix83-85, que se asocia a mayor tasa de fracaso a la RT, QT y al aumento de las metástasis66.
Sabemos que la activación de la glucólisis aeróbica representa un evento inicial del proceso de transformación neoplásica, probablemente como respuesta al aumento de la proliferación celular86, ya que las células con una elevada tasa de proliferación consumen más oxígeno. Los tumores tienen un aumento de la glucólisis y sabemos que la concentración de la glucosa y de los componentes de la vía glucolítica son influyentes en el HIF87,88. El pH tumoral es más ácido debido a la mayor producción de lactato y CO2. Para sobrevivir, las células necesitan mantener un equilibrio entre el pH intracelular y el extracelular; esto se logra a costa de varios transportadores. La anhidrasa carbónica IX (CA9) ejerce un papel fundamental en este equilibrio; varios estudios han demostrado la correlación entre la hipoxia, angiogénesis, HIF-1a y el CA989.
Por tanto, los niveles del HIF son adaptados para que las células mantengan una elevada tasa de proliferación y, por otra parte, el aumento de la proliferación celular puede inducir una mayor expresión del HIF28. Ante las situaciones de hipoxia, en las que la acción de los factores de crecimiento lleven a un aumento de la proliferación celular y, por tanto, a una mayor necesidad de oxígeno, el HIF-1a será más expresado y activado, induciendo a la expresión de genes que codifican las moléculas proangiogénicas y que permiten una adaptación metabólica a la hipoxia, siendo el activador más potente de genes que codifican las enzimas glucolíticas y los factores de crecimiento proangiogénicos28,90-93, ya que los tumores no pueden progresar sin la angiogénesis que permite la difusión del oxígeno, la glucosa y otros nutrientes77,78.
La angiogénesis consiste en el desarrollo de nuevos vasos a partir de la red vascular preexistente y tiene un papel preponderante en los diversos mecanismos fisiopatológicos benignos (cicatrización, heridas, isquemia, retinopatía diabética) y malignos (el crecimiento del tumor y metástasis); el VEGF desempeña un papel fundamental en la angiogénesis y está regulado por el HIF94-96.
Actualmente, existen evidencias de que los vasos tumorales son desorganizados y sin estructura adecuada para la circulación, lo que conduce frecuentemente al colapso. Dado que el crecimiento tumoral requiere oxígeno, nutrientes y una función metabólica apropiada para su desarrollo, es necesario promover los factores de la angiogénesis para inhibir la apoptosis de las células tumorales desencadenadas por la hipoxia. Por tanto, la angiogénesis como respuesta a la hipoxia tumoral está mediada por el HIF-1a55.
El HIF-1a se ha considerado un factor clave en la regulación del VEGF y su receptor (VEGRF), como de otros factores angiogénicos. Varios estudios inmunohistoquímicos realizados en diversos modelos tumorales71 revelan que la expresión del HIF-1a está asociada a un aumento del VEGF y de la vascularización y metastización, que implican un peor pronóstico72,76. Parecer existir una relación directa entre la angiogénesis y la metastización en diversos tipos de tumores, como melanomas, gliomas, pulmón, mama, ovario vejiga y próstata97,98, comprobándose que las proteínas diana del HIF-1a están implicadas en la proliferación, supervivencia, adhesión y movilidad de las células neoplásicas.
Por otra parte, el aumento de la expresión del HIF-1a, combinada con las mutaciones inactivadas en los genes supresores como VHL, p53, PTEN o la amplificación de los oncogenes Akt, RAS, ERK1/2, se ha observado con frecuencia en los pacientes oncológicos; estas alteraciones están asociadas a crecimiento tumoral, invasión y metástasis.
Zhong et al99 han demostrado una expresión aumentada del HIF-1a en alrededor del 53% de los tumores, incluidos el de colon, gástricos, pancreáticos, pulmón, ovario, próstata, renal, melanoma y glioblastoma. El aumento de expresión del HIF-1a está asociado a una supervivencia más corta en el cáncer de mama y de útero, y a la mala respuesta al tratamiento en el cáncer nasofaríngeo, resaltando el papel de la hipoxia tumoral en el pronóstico72,100-104 (tabla 2).

En el cáncer de próstata, se encuentra expresado en los estadios iniciales de la carcinogénesis y esa expresión está relacionada con los indicadores de diagnóstico y pronóstico para la rediviva temprana y metastización, pudiendo ser el HIF-1 un potencial biomarcador del pronóstico. Su importancia en la progresión tumoral lo convierte en un objetivo plausible en las estrategias de quimioprevención, así como en la capacidad de inhibir la angiogénesis60. Los estudios experimentales realizados en células de cáncer de próstata en ratones muestran que una sobreexpresión del HIF-1a se asocia a un mayor crecimiento y potencial metastático108. Del mismo modo, en los seres humanos también se ha encontrado una mayor expresión del HIF-1a en los tumores de próstata48,99. En el CaP, en particular los pacientes con cáncer metastizado u hormonorresistente, se ha demostrado que el gen VEGF, principalmente inducido por el HIF-1a está frecuentemente sobreexpresado, lo que sugiere una acción central de esta molécula en este proceso105,106.
La activación de los oncogenes y los factores de crecimiento pueden inducir el sistema HIF en las células no hipoxidantes o amplificar la respuesta a la hipoxia. De hecho, varios factores de crecimiento y citocinas del estroma y parénquima también actúan como reguladores y son capaces de inducir la expresión del HIF-1a, su capacidad de enlace y de transactivación, como por ejemplo el factor de crecimiento epidérmico (EGF)46, el TGFa92,107 los factores IGF-1 e IGF-2109 y la interleucina 1b110. Además, estudios recientes muestran que el HIF puede desempeñar un papel importante en la resistencia a los tratamientos111-113.
El sistema HIF actúa como el máximo regulador de la respuesta a la hipoxia, iniciando la cascada de mecanismos que permiten que el tumor se adapte al medio ambiente hostil, y surge como un importante factor de transcripción en la biología del cáncer.
Conclusión
La activación del HIF es regulada por diversos mecanismos a partir de la estabilización de la subunidad HIF-1a que envuelve múltiples señales y vías.
La hipoxia, algunos genes supresores tumorales, factores de crecimiento y citocinas aumentarán la estabilidad y/o transactivación del HIF1A, lo que resulta en un incremento de la producción del HIF-1a y, como consecuencia, la angiogénesis tumoral, la adaptación metabólica a la hipoxia y el aumento de la supervivencia celular, debido a su acción en varios genes diana. El HIF-1a será crucial en la iniciación de la angiogénesis, en el crecimiento tumoral, la progresión y la metastización.
De esta forma, nos parece fundamental desenvolver técnicas para bloquear o inhibir la angiogénesis y el factor HIF1a, reduciendo la posibilidad de desarrollar neoplasias más agresivas y, de este modo, reducir la morbilidad y mortalidad por cáncer.
El HIF-1a podría constituir un marcador temprano de la carcinogénesis, con valor predictivo de progresión tumoral y valor pronóstico.

Figura 6 - Papel del HIF en la supervivencia y la muerte celular.
Adaptado de Acker y Plate93.
Bibliografía
1. Semenza GL. HIF-1 and human disease: One highly involved factor. Genes Dev. 2000;14:1983-91. [ Links ]
2. Semenza GL. Targeting HIF-1 for cancer therapy. Nat Rev Cancer. 2003;3:721-32. [ Links ]
3. Kaelin WG Jr. Molecular basis of the VHL hereditary cancer syndrome. Nat Rev Cancer. 2002;2:673-82. [ Links ]
4. Wang GL, Semenza GL. Purification and characterization of hypoxia-inducible factor 1. J Biol Chem. 1995;270:1230-7. [ Links ]
5. Semenza GL. HIF 1 and tumor progression: pathophysiology and therapeutics. Trends Mol Med. 2002;8:S62-7. [ Links ]
6. Semenza GL. Involvement of HIF 1 in human cancer. Intern Med. 2002;41:79-83. [ Links ]
7. Manalo DJ, Rowan A, Lavoie T, Natarajan L, Kelly BD, Ye SQ, et al. Transcriptional regulation of vascular endothelial cell responses to hypoxia by HIF-1. Blood. 2005;105:659-69. [ Links ]
8. Jain RK. Normalizing tumor vasculature with anti angiogenic therapy: a new paradigm for combination therapy. Nat Med. 2001;7:987-9. [ Links ]
9. Semenza GL, Rue EA, Iyer NV, Pang MG, Kearns WG. Assignment of the hypoxia inducible factor 1a gene to a region of conserved synteny on mouse chromosome 12 and human chromosome 14q. Genomics. 1996;34:437-9. [ Links ]
10. Iyer NV, Leung SW, Semenza GL. The human hypoxia inducible factor 1 alpha gene: HIF1A structure and evolutionary conservation. Genomics. 1998;52:159-65. [ Links ]
11. Tian H, McKnight SL, Russell DW. Endothelial PAS domain protein 1 (EPAS1), a transcription factor selectively expressed in endothelial cells. Genes Dev. 1997;11:72-82. [ Links ]
12. Ema M, Taya S, Yokotani N, Sogawa K, Matsuda Y, Fujii-Kuriyama Y. A novel bHLH-PAS factor with close sequence similarity to hypoxia-inducible factor 1alpha regulates the VEGF expression and is potentially involved in lung and vascular development. Proc Natl Acad Sci USA. 1997;94:4273-8. [ Links ]
13. Flamme I, Frohlich T, Von Reutern M, Kappel A, Damert A, Risau W. HRF, a putative basic helix-loop-helix-PAS-domain transcription factor is closely related to hypoxia-inducible factor-1 alpha and developmentally expressed in blood vessels. Mech Dev. 1997;63:51-60. [ Links ]
14. Gu YZ, Moran SM, Hogenesch JB, Wartman L, Bradfield CA. Molecular characterization and chromosomal localization of a third alpha-class hypoxia inducible factor subunit, HIF3alpha. Gene Expr. 1998;7:205-13. [ Links ]
15. Wenger RH. Cellular adaptation to hypoxia: O2-sensing protein hydroxylases, hypoxia-inducible transcription factores, and O2-regulated gene expression. FASEB J 2002;16:1151-62. [ Links ]
16. Vandromme M, GauthierRouviere C, Lamb N, Fernández A. Regulation of transcription factor localization: Fine-tuning of gene expression. Trends Biochem Sci. 1996;21:59-64. [ Links ]
17. Jiang BH, Zheng JZ, Leung SW, Roe R, Semenza GL. Transactivation and inhibitory domains of hypoxiainducible factor 1 alpha. Modulation of transcriptional activity by oxygen tension. J Biol Chem. 1997;272:19253-60. [ Links ]
18. Jiang BH, Rue E, Wang GL, Roe R, Semenza GL. Dimerization, DNA binding, and transactivation properties of hypoxiainducible factor 1. J Biol Chem. 1996;271:17771-8. [ Links ]
19. Michel G, Minet E, Ernest I, Roland I, Durant F, Remacle J, et al. A model for the complex between the hypoxiainducible factor-1 (HIF-1) and its consensus DNA sequence. J Biomol Struc Dyn. 2000;18:169-79. [ Links ]
20. Huang LE, Gu J, Schau M, Bunn HF. Regulation of hypoxiainducible factor 1 alpha is mediated by an O-2-dependent degradation domain via the ubiquitin- proteasome pathway. Proc Natl Acad Sci USA. 1998;95:7987-92. [ Links ]
21. Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, et al. HIFalpha targeted for VHLmediated destruction by proline hydroxylation: Implications for O2 sensing. Science. 2001;292:464-8. [ Links ]
22. Bruick RK, McKnight SL. A conserved family of prolyl-4-hydroxylases that modify HIF. Science. 2001;294:1337-40. [ Links ]
23. Pause A, Lee S, Worrell RA, Chen DYT, Burguess WH, Marston Linehan W, et al. The von Hipple-Lindau tumor suppressor gene product forms a stable complex with human cul-2, a member of the Cdc53 family of proteins. Proc Natl Acad Sci USA. 1997;94:2156-61. [ Links ]
24. Lonergan KM, Iliopoulos O, Ohh M, Kamura T, Conaway RC, Conaway JW, et al. Regulation of hypoxia ¿inducible mRNAs by the von Hipple-Lindau tumor suppressor protein requires binding to complexes containing elongins B/C and Cul2. Mol Cell Biol. 1998;18:732-41. [ Links ]
25. Maxwell PH, Wiesener MS, Chang GW, Clifford SC, Vaux EC, Cockman ME, et al. The tumor suppressor protein VHL targets hypoxia inducible factores for oxygen -dependent proteolysis. Nature. 1999;399:271-5. [ Links ]
26. Ohh M, Park CW, Ivan M, Hoffman MA, Kim TY, Huang LE, et al. Ubiquitination of hypoxia inducible factor requires direct binding to the beta domain of the von Hipple-Lindau protein. Nat Cell Biol 2000;2:423-7. [ Links ]
27. Tanimoto K, Makino Y, Pereira T, Poellinger L. Mechanism of regulation of the hypoxia inducible factor 1 alpha by the von Hipple Lindau tumor suppressor protein. EMBO J. 2000;19:4298-309. [ Links ]
28. Semenza GL, Jiang BH, Leung SW, Passantino R, Concordet JP, Maire P, et al. Hypoxia response elements in the aldolase A, enolase 1, and lactate dehydrogenase A gene promoters contain essential binding sites for hypoxia- inducible factor 1. J Biol Chem. 1996;271:32529-37. [ Links ]
29. Epstein AC, Gleadle JM, McNeill LA, Hewitson KS, O´Rourke J, Mole DR, et al. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell.2001;107:43-54. [ Links ]
30. Min JH, Yang H, Ivan M, Gertler F, Kaelin WG Jr, Pavletich NP. Structure of an HIF-1alpha-pVHL complex: Hydroxyproline recognition in signaling. Science. 2002;296:1886-9. [ Links ]
31. Yu F, White SB, Zhao Q, Lee FS. HIF-1alpha binding to VHL is regulated by stimulus-sensitive proline hydroxylation. Proc Natl Acad Sci USA. 2001;98:9630-5. [ Links ]
32. Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, et al. Targeting of HIF-alpha to the von Hippel Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science. 2001;292:468-72. [ Links ]
33. Masson N, Willam C, Maxwell PH, Pugh CW, Ratcliffe PJ. Independent function of two destruction domains in hypoxiainducible factor-alpha chains activated by prolyl hydroxylation. EMBO J. 2001;20:5197-6. [ Links ]
34. Jeong JW, Bae MK, Ahn MY, Kim SH, Sohn TK, Bae MH, et al. Regulation and destabilization of HIF-1alpha by ARD1-mediated acetylation. Cell. 2002;111:709-20. [ Links ]
35. Lando D, Peet DJ, Whelan DA, Gorman JJ, Whitelaw ML. Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch. Science. 2002;295:858-61. [ Links ]
36. Boddy JL. The androgen receptor is significantly associated with vascular endothelial growth factor and hypoxia sensing via HIF1a, HIF2a and the prolyl hydroxylases in human prostate cancer. Clin Cancer Res. 2005;11.7658-63. [ Links ]
37. Wykoff CC, Sotiriou C, Cockman ME, Ratcliffe PJ; Maxwell P, Liu E, et al. Gene array of VHL mutation and hipoxia shows novel hipoxia induced genes and that cyclin D1 is a VHL target gene. Br J Cancer. 2004;90:1235-43. [ Links ]
38. Melillo G. Inhibiting hypoxia inducible factor 1 for cancer therapy. Mol Cancer Res. 2006;4:601-5. [ Links ]
39. Chan DA, Sutphin PD, Denko NC, Giaccia AJ. Role of prolyl hydroxylation in oncogenically stabilized hypoxia-inducible factor-1alpha. J Biol Chem. 2002;277:40112-7. [ Links ]
40. Dameron KM, Volpert OV, Tainsky MA, Bouck N. Control of angiogenesis in fibroblasts by P53 regulation of thrombospondin-1. Science. 1994;265:1582-4. [ Links ]
41. Bouvet M, Ellis LM, Nishizaki M, Fujiwara T, Liu WB, Bucana CD, et al. Adenovirus-mediated wild-type p53 gene transfer down-regulates vascular endothelial growth factor expression and inhibits angiogenesis in human colon cancer. Cancer Res. 1998;58:2288-92. [ Links ]
42. Blancher C, Moore JW, Robertson N, Harris AL. Effects of ras and von Hippel-Lindau (VHL) gene mutations on hypoxiainducible factor (HIF)-1 alpha, HIF-2 alpha, and vascular endothelial growth factor expression and their regulation by the phosphatidylinositol 3 ´-kinase/Akt signaling pathway. Cancer Res. 2001;61:7349-55. [ Links ]
43. Laughner E, Taghavi P, Chiles K, Mahon PC, Semenza GL. HER2 (neu) signaling increases the rate of hypoxiainducible factor 1alpha (HIF-1alpha) synthesis: Novel mechanism for HIF-1-mediated vascular endothelial growth factor expression. Mol Cell Biol. 2001;21:3995-4004. [ Links ]
44. Arsham AM, Plas DR, Thompson CB, Simon MC. Phosphatidylinositol 3-kinase/Akt signaling is neither required for hypoxic stabilization of HIF-1 alpha nor sufficient for HIF-1-dependent target gene transcription. J Biol Chem. 2002;277:15162-70. [ Links ]
45. Mottet D, Dumont V, Deccache Y, Demazy C, Ninane N, Raes M, et al. Regulation of hypoxia-inducible factor-1alpha protein level during hypoxic conditions by the phosphatidylinositol 3-kinase/Akt/glycogen synthase kinase 3beta pathway in HepG2 cells. J Biol Chem. 2003;278:31277-85. [ Links ]
46. Zhong H, Chiles K, Feldser D, Laughner E, Hanrahan C, Georgescu MM, et al. Modulation of hypoxiainducible factor 1 alpha expression by the epidermal growth factor/phosphatidylinositol 3- inase/PTEN/AKT/FRAP pathway in human prostate cancer cells: Implications for tumor angiogenesis and therapeutics. Cancer Res. 2000;60:1541-5. [ Links ]
47. Mabjeesh NJ, Willard MT, Frederickson CE, Zhong H, Simons JW. Androgens stimulate hypoxia-inducible factor 1 activation via autocrine loop of tyrosine kinase receptor/phosphatidylinositol 3´-kinase/protein kinase B in prostate cancer cells. Clin Cancer Res. 2003;9:2416-25. [ Links ]
48. Outi RS, Kimmo JS, Nina NN, Ola B, Tapio V. Amplification of HIF 1a gene in prostate cancer. Cancer Genet Cytogenet. 2001;128:31-4. [ Links ]
49. Gao N, Ding M, Zheng JZ, Shi X, Jiang BH. Vanadate-induced expression of hypoxia-inducible factor 1 alpha and vascular endothelial growth factor through phosphatidylinositol 3-kinase/Akt pathway and reactive oxygen species. J Biol Chem. 2002;277:31963-71. [ Links ]
50. Gao N, Jiang BH, Corum L, Roberts JR, Antonini J, Zheng JZ, et al. p38 aignaling-mediated hypoxia- inducible factor 1alpha and vascular endothelial growth factor induction by Cr(VI) in DU145 human prostate carcinoma cells. J Biol Chem. 2002;277:45041-8. [ Links ]
51. Haddad JJ, Land SC. A non-hypoxic, ROS-sensitive pathway mediates TNF-alpha-dependent regulation of HIF-1alpha. FEBS Lett. 2001;505:269-74. [ Links ]
52. Chandel NS, McClintock DS, Feliciano CE, Wood TM, Melendez JA, Rodriguez AM, et al. Reactive oxygen species generated at mitochondrial complex III stabilize hypoxiainducible factor-1 alpha during hypoxia - A mechanism of O-2 sensing. J Biol Chem. 2000;275:25130-8. [ Links ]
53. Zander R, Vaupel P. Proposal for using a standardized terminology on oxygen transport to tissue. Adv Exp Med Biol. 1985;191:965-70. [ Links ]
54. Stroka DM, Burkhardt T, Desbaillets I, Wenger RH, Neil DA, Bauer C, et al. HIF-1 is expressed in normoxic tissue and displays an organ-specific regulation under systemic hypoxia. FASEB J. 2001;15:2445-53. [ Links ]
55. Yong-HS, Wei GF. Hypoxia-inducible factor-1 in tumor angiogenesis. World J Gastroenterol. 2004;10:1082-7. [ Links ]
56. Vaupel P, Kallinowski F, Okunieff P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: a review. Cancer Res. 1989;49:6449-65. [ Links ]
57. Kimura H, Braun RD, Ong ET, Hsu R, Secomb TW, Papahadjopoulos D, et al. Fluctuations in red cell flux in tumor microvessels can lead to transient hypoxia and reoxygenation in tumor parenchyma. Cancer Res. 1996;56:5522-8. [ Links ]
58. Brown JM, Giaccia AJ. The unique physiology of solid tumors: opportunities (and problems) for cancer therapy. Cancer Res. 1998;58:1408-16. [ Links ]
59. Dewhirst MW. Concepts of oxygen transport at the microcirculatory level. Semin Radiat Oncol. 1998;8:143-50. [ Links ]
60. Kimbro KS, Simons JW. Hypoxia-inducible factor in human breast and prostate cancer. Endocrine Related Cancer. 2006;13:739-49. [ Links ]
61. Kaelin WG. ROS: really involved in oxygen sensing. Cell Metabolism. 2005;1:357-8. [ Links ]
62. Muzandu K, Shaban Z, Ishizuka M, Kasusaka A, Fujita S. Nitric Oxide enhances catechol estrogen induced oxidative stress in LNCaP cells. Free Radical Research. 2005;39:389-98. [ Links ]
63. Semenza GL. Regulation of mammalian O2 homeostasis by hypoxia inducible factor 1. Annu Ver Cell Dev Biol. 1999;15:551-78. [ Links ]
64. Gleadle JM, Ratcliffe PJ. Induction of hypoxia inducible factor 1, erythropoietin, vascular endothelial growth factor and glucose transporter 1 by hypoxia: evidence against a regulatory role for Src kinase. Blood. 1997;89:503-9. [ Links ]
65. Airley R, Loncaster J, Davidson S, Bromley M, Roberts S, Patterson A, et al. Glucose transporter glut 1 expression correlates with tumor hypoxia and predicts metastasis free survival in advanced carcinoma of the cervix. Clin Cancer Res. 2001;7:928-34. [ Links ]
66. Bristow RG, Richard P. Hypoxia, DNA repair and genetic instability. Hill in Nature Reviews. 2008;8:180-92. [ Links ]
67. Prince BD, Calderwood SK. GADD45 and GADD153 messenger RNA levels are increased during hypoxia and after exposure of cells to agents which elevate the levels of the glucose regulated proteins. Cancer Res. 1992;52:3814-7. [ Links ]
68. Semenza GL. Angiogenesis in ischemic and neoplastic disorders. Annu Rev Med. 2003;54:17-28. [ Links ]
69. Dachs GU, Tozer GM. Hypoxia modulated gene expression: angiogenesis, metastasis and therapeutic exploitation. Eur J Cancer. 2000;36:1649-60. [ Links ]
70. Forsythe JA, Jiang BH, Iyer NV, Agani F, Leung SW, Koods RD, et al. Activation of vascular endothelial growth factor gene transcription by hypoxia inducible factor 1. Mol Cell Biol. 1996;16:4604-13. [ Links ]
71. Talks KL, Turley H, Gatter KC, Maxwell PH, Pugh CW, Ratcliffe PJ, et al. The expression and distribution of the hypoxiainducible factors HIF-1 alpha and HIF-2 alpha in normal human tissues, cancers, and tumor-associated macrophages. Am J Pathol. 2000;157:411-21. [ Links ]
72. Bos R, Zhong H, Hanrahan CF, Mommers EC, Semenza GL, Pinedo HM, et al. Levels of hypoxia-inducible factor-1 alpha during breast carcinogenesis. J Natl Cancer Inst. 2001;93:309-14. [ Links ]
73. Giatromanolaki A, Koukourakis MI, Sivridis E, Turley H, Talks K, Pezzella F, et al. Relation of hypoxia inducible factor 1 alpha and 2 alpha in operable non-small cell lung cancer to angiogenic/molecular profile of tumours and survival. Br J Cancer. 2001;85:881-90. [ Links ]
74. Shi BM, Wang XY, Mu QL, Wu TH, Liu HJ, Yang Z. Angiogenesis effect on rat liver after administration of expression vector encoding vascular endothelial growth factor D. World J Gastroenterol. 2003;9:312-5. [ Links ]
75. Toi M, Hoshina S, Takayangi T, Tominaga T. Association of vascular endothelial growth-factor expression with tumor angiogenesis and with early relapse in primary breastcancer. Jpn Cancer Res. 1994;85:1045-9. [ Links ]
76. Maeda K, Chung YS, Ogawa Y, Takatsuka S, Kang SM, Ogawa M, et al. Prognostic value of vascular endothelial growth factor expression in gastric carcinoma. Cancer. 1996;77:858-63. [ Links ]
77. Huss WJ, Hanrahan CF, Barrios RJ, Simons JW, Greenberg NM. Angiogenesis and prostate cancer: Identification of a molecular progression switch. Cancer Res. 2001;61:2736-43. [ Links ]
78. Kerbel RS. New targets, drugs and approaches for the treatment of cancer: an overview. Cancer Metastasis Rev. 1998;17:145-7. [ Links ]
79. Vaupel P. The role of hypoxia induced factores in tumor progression. Oncologist. 2004;9 Suppl 5:10-7. [ Links ]
80. Vaupel P, Kelleher DK, Hockel M. Oxygen status of malignant tumors: pathogenesis of hypoxia and significance for tumor therapy. Seminars in Oncology. 2001;28:29-35. [ Links ]
81. Acs G, Xu X, Chu C, Acs P, Verma A. Prognostic significance of erythropoietin expression in human endometrial carcinoma. Cancer. 2004;100:2376-86. [ Links ]
82. Vleugel MM, Greijer AE, Shvarts A, Van Der Groep P, Van Berkel M, Aarbodem Y, Van Tinteren H, et al. Differential prognostic impact of hypoxia induced and diffuse HIF-1alpha expression in invasive breast cancer. J Clin Pathol. 2005;58:172-7. [ Links ]
83. Chan N, Milosevic M, Bristow RG. Tumor hypoxia, DNA repair and prostate cancer progression: new targets and new therapies. Future Oncol. 2007;3:329-41. [ Links ]
84. Chaudary N, Hill RP. Hypoxia and metastasis. Clin Cancer Res. 2007;13:1947-9. [ Links ]
85. Vaupel P, Mayer A. Hypoxia in cancer: significance and impact in clinical outcome. Cancer Metastasis Ver. 2007;26:225-39. [ Links ]
86. Brand KA, Hermfisse U. Aerobic glycolysis by proliferating cells: a protective strategy against reactive oxygen species. FASEB J. 1997;11:388-95. [ Links ]
87. Gatenby RA, Gilles RJ. Why do cancers have high aerobic glycolysis? Nat Rev Cancer. 2004;4:891-9. [ Links ]
88. Vordermark D, Kraft P, Katzer A, Bolling T, Willner J, Flentje M. Glucose requirement for hypoxic accumulation of hypoxia inducible factor 1alpha. Cancer Lett. 2005;230:122-33. [ Links ]
89. Van den Eynden GG, Van der Auwera I, Van Laere SJ, Colpaert CG, Turley H, Harris AL, et al. Angiogenesis and hypoxia in lymph node metastasis is predicted by the angiogenesis and hypoxia in the primary tumour in patients with breast cancer. BR J Cancer. 2005;93:1128-36. [ Links ]
90. Semenza GL. HIF-1: mediator of physiological and pathophysiological responses to hypoxia. J Appl Physiol. 2000;88:1474-80. [ Links ]
91. Ryan HE, Lo J, Johnson RS. HIF-1 alpha is required for solid tumor formation and embryonic vascularization. Embol J. 1998;17:3005-15. [ Links ]
92. Acker T, Plate KH. A role of hypoxia and hypoxia inducible transcription factores in tumor physiology. J Mol Med. 2002;80:562-75. [ Links ]
93. Acker T, Plate KH. A role of hypoxia in tumor angiogenesis-molecular and cellular angiogenic crosstalk. Cell Tissue Res. 2003;80:562-75. [ Links ]
94. Brahimi-Horn MC, Pouyssegur J. The hypoxia inducible factor and tumor progression along the angiogenic pathway. Int Rev Cytol. 2005;242:157-213. [ Links ]
95. Pugh CW, Ratcliffe PJ. Regulation of angiogenesis by hypoxia: role of the HIF system. Nat Med. 2003;9:677-84. [ Links ]
96. Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and its receptores. Nat Med. 2003;9:669-76. [ Links ]
97. Bochner BH, Cote RJ, Weidner N, Groshen S, Chen SC, Skinner DG, et al. Angiogenesis in bladder cancer relationship between microvessel density and tumor prognosis. J Natl Cancer Inst. 1995;87:1603-12. [ Links ]
98. Jaeger TM, Weidner N, Chew K, Moore DH, Kerschmann RL, Waldman FM, et al. Tumor angiogenesis correlateswith lymph-node metastases in invasive bladder-cancer. J Urol. 1995;154:69-71. [ Links ]
99. Zhong H, De Marzo AM, Laughner E, Lim M, Hilton DA, Zagzag D, et al. Overexpression of hypoxia-inducible factor 1alpha in common human cancers and their metastases. Cancer Res. 1999;59:5830-5. [ Links ]
100. Bos R, Van der Greijer AE, Shvarts A, Meijer S, Pinedo HM, Semenza GL, et al. Levels of hypoxiainducible factor-1alpha independently predict prognosis in patients with lymph node negative breast carcinoma. Cancer. 2003;97:1573-81. [ Links ]
101. Birner P, Schindl M, Obermair A, Breitenecker G, Oberhuber G. Expression of hypoxia-inducible factor 1alpha in epithelial ovarian tumors: its impact on prognosis and on response to chemotherapy. Clin Cancer Res. 2001;7:1661-8. [ Links ]
102. Aebersold DM, Burri P, Beer KT, Laissue J, Djonov V, Greiner RH, et al. Expression of hypoxia-inducible factor-1alpha: a novel predictive and prognostic parameter in the radiotherapy of oropharyngeal cancer. Cancer Res. 2001;61:2911-6. [ Links ]
103. Hockel M, Vaupel P. Tumor hypoxia: definitions and current clinical, biologic, and molecular aspects. J Natl Cancer Inst. 2001;93:266-76. [ Links ]
104. Li Z, Wang D, Na X, Schoen SR, Messing EM, Wu G. The VHL protein recruits a novel KRAB-A domain protein to repress HIF-1alpha transcriptional activity. EMBO J. 2003;22:1857-67. [ Links ]
105. Kondo K, Klco J, Nakamura E, Lechpammer M, Kaelin WG Jr. Inhibition of HIF is necessary for tumor suppression by the von Hippel-Lindau protein. Cancer Cell. 2002;1:237¿46. [ Links ]
106. George DJ, Regan MM, Oh WK, Tay MH, Manola J, Decalo N, et al. Radical prostatectomy lowers plasma vascular endothelial growth factor levels in patients with prostate cancer. Urology. 2004;63:327-32. [ Links ]
107. Krishnamachary B, Berg-Dixon S, Kelly B, Agani F, Feldser D, Ferreira G, et al. Regulation of colon carcinoma cell invasion by hypoxia-inducible factor 1. Cancer Res. 2003;63:1138-43. [ Links ]
108. Zhong H, Agani F, Baccala AA, Laughner E, Rioseco-Camacho N, Isaacs WB, et al. Increased expression of hypoxia inducible factor 1 alpha in rat and human prostate cancer. Cancer Res. 1998; 58:5280-4. [ Links ]
109. Treins C, Giorgetti-Peraldi S, Murdaca J, Semenza GL, Van Obberghen E. Insulin stimulates hypoxia-inducible factor 1 through a phosphatidylinositol 3-kinase/target of rapamycindependent signaling pathway. J Biol Chem. 2002;277:27975-81. [ Links ]
110. Stiehl DP, Jelkmann W, Wenger RH, Hellwig-Burgel T. Normoxic induction of the hypoxia-inducible factor 1alpha by insulin and interleukin-1beta involves the phosphatidylinositol 3-kinase pathway. FEBS Lett. 2002;512:157-62. [ Links ]
111. Brown JM, Giaccia AJ. The unique physiology of solid tumors: opportunities and problems for cancer therapy. Cancer Res. 1998;58:1408-16. [ Links ]
112. Wartenberg M, Ling FC, Muschen M, Klein F, Acker H, Gassmann M, et al. Regulation of the multidrug resistance transporter P-glycoprotein in multicellular tumor spheroids by hypoxia-inducible factor (HIF-1) and reactive oxygen species. FASEB J. 2003;17:503-5. [ Links ]
113. Comerford KM, Wallace TJ, Karhausen J, Louis NA, Montalto MC, Colgan SP, et al. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res. 2002;62:3387-94. [ Links ]
![]() Dirección para correspondencia:
Dirección para correspondencia:
Correo electrónico: avfraga@gmail.com
(A. Fraga).
Recibido: 16 de abril de 2009
Aceptado: 25 de junio de 2009

