










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkNutrición Hospitalaria
versión On-line ISSN 1699-5198versión impresa ISSN 0212-1611
Nutr. Hosp. vol.27 no.6 Madrid nov./dic. 2012
https://dx.doi.org/10.3305/nh.2012.27.6.6140
Nootropic and hypophagic effects following long term intake of almonds (Prunus amygdalus) in rats
Efectos nootróficos e hipofágicos del consumo prolongado de almendras (Prunus amygdalus) en ratas
S. Haider1, Z. Batool1 and D. J. Haleem1,2
1Neurochemistry and Biochemical Neuropharmacology Research Unit. Department of Biochemistry. University of Karachi. Karachi. Pakistan
2Neuroscience Research Library. Dr. Panjwani Center for Molecular Medicine and Drug Research. University of Karachi. Pakistan
The authors are thankful to the University of Karachi, Pakistan for funding this project.
ABSTRACT
Introduction: Over a period of time researchers have become more interested in finding out the potential of various foods to maintain the general health and to treat diseases. Almonds are a very good source of many nutrients which may help to sharpen the memory and to reduce cardiovascular risk factors.
Objective: The present study was conducted to evaluate the nootropic effects of almonds. Effect of oral intake of almond was also monitored on food intake and plasma cholesterol levels.
Methods: Rats were given almond paste orally with the help of feeding tube for 28 days. Memory function in rats was assessed by Elevated Plus Maze (EPM) and Radial Arm Maze (RAM). Brain tryptophan, 5-HT and 5-HIAA were estimated at the end of the treatment by HPLC-EC method.
Results: A significant improvement in learning and memory of almond treated rats compared to controls was observed. Almond treated rats also exhibited a significant decrease in food intake and plasma cholesterol levels while the change in growth rate (in terms of percentage) remained comparable between the two groups. Analysis of brain tryptophan (TRP) monoamines exhibited enhanced TRP levels and serotonergic turnover in rat brain following oral intake of almonds.
Conclusion: The findings show that almonds possess significant hypophagic and nootropic effects. Results are discussed in context of enhanced 5-HT metabolism following almond administration.
Key words: 5-HT turnover. Almond. Cholesterol. Hypophagic action. Nootropic substance.
RESUMEN
Introducción: De un tiempo a esta parte, los investigadores se han interesado cada vez más en encontrar el potencial de diversos alimentos para mantener la salud en general y tratar la enfermedad. Las almendras son una muy buen fuente de muchos nutrientes que pueden ayudar a agudizar la memoria y reducir los factores de riesgo cardiovascular.
Objetivo: Este estudio se realizó para evaluar los efectos nootróficos de la almendra. También se monitoreó el efecto de la ingestión oral de almendra sobre el consumo de alimentos y las concentraciones plasmáticas de colesterol.
Métodos: Se suministró a las ratas pasta de almendra por vía oral con la ayuda de una sonda de alimentación durante 28 días. Se evaluó la función de memoria de las ratas mediante Elevated Plus Maze (EPM) y Radial Arm Maze (RAM). Se estimaron el triptófano cerebral, la 5-HT y el 5-HIAA al final del tratamiento mediante un método de HPLC-EC.
Resultados: Se observó una mejoría significativa en el aprendizaje y la memoria en las ratas tratadas con almendra en comparación con los controles. Las ratas tratadas con almendra también mostraron una reducción significativa del consumo de alimentos y de las concentraciones plasmáticas de colesterol, mientras que el cambio en la tasa de crecimiento (en términos de porcentaje) siguió siendo comparable entre ambos grupos. El análisis de las monomaninas y de triptófano cerebral mostró unas concentraciones superiores de triptófano y del recambio serotoninérgico del cerebro de la rata tras el consumo de almendras.
Conclusión: los hallazgos sugieren que las almendras poseen unos efectos hipofágicos y nootróficos significativos. Los resultados se debaten en el contexto del metabolismo mejorado de la 5-HT tras la administración de almendras.
Palabras clave: Recambio de 5-HT. Almendra. Colesterol. Acción hipofágica. Sustancia nootrófica.
Abbreviations
5-HIAA: 5-hydroxy indole acetic acid.
5-HT: 5-hydroxy tryptamine.
CVD: Cardiovascular disease.
EPM: Elevated plus maze.
HPLC-EC: High performance liquid chromatography with electrochemical detection.
PUFAs: Polyunsaturated fatty acids.
RAM: Radial arm maze.
TRP: Tryptophan.
Introduction
Diet plays an influential role in the maintenance of general health as well as in the prevention and treatment of diseases. Memory loss1 and obesity2 are two of the prevalent diseases throughout the world. With the passage of time there is increased trend to maintain general health through diet. Various foods including tree nuts have been shown to affect central nervous system,3 enhance learning, memory and intelligence.4 Memory formation is the complex process and involves various neurotransmitters and neuronal pathways.5 It is well established that serotonergic system has an important role in memory function.6 Tryptophan (TRP), the precursor of serotonin (5-hydroxy tryptamine; 5HT) and an essential amino acid, has also been shown to increase learning and memory.7,8 Source of TRP is dietary only and increased brain TRP availability has been revealed to increase brain serotonin synthesis.9
Obesity and being overweight are linked to reduce quality of life, greater morbidity10 and mortality.11 Elevated cholesterol as a result of obesity is one of the risk factor for cardiovascular diseases (CVD).12 Nuts are high-fat, energy rich foods (e.g. pecan 70%, macadamia nut 66%, Brazil nut 65%, walnut 60%, almonds 55%),13 so it is logical that frequent consumption of nuts may lead to increased cholesterol and weight gain and, consequently, increase the risk of cardiovascular disease and many chronic diseases. But prior studies did not show any adverse effect of nut consumption on energy balance or body weight. 14 In fact, a growing body of evidence indicates that eating nuts on the regular basis may be linked to decreased weight gain as well as cholesterol levels in various human populations. 15-17
Nuts are energy-dense and nutrients loaded foods. Walnuts and almonds both have been used as a nootropic substance from time immemorial. To experimentally show this we previously analyzed the walnut.18 The findings tempted us to investigate the effects of almonds on learning and memory functions. These nuts contain a wide variety of bioactive compounds that seem to have constructive effects on human health.19 According to US department of Agriculture Nutrient Database, almonds have a high protein content (21.22 g/100 g of raw almond) and the TRP content of this nut is about 214 mg/100 g of almond. These nuts provide 5.75 kcal/g of energy and are low in saturated fat (< 8% of total fats) but rich in unsaturated fats (92% of total fats), mainly of which are monounsaturated (66.15%). Almonds are one of the most important source of plant protein, and good source of dietary fiber, minerals, antioxidants, vitamins and numerous bioactive substances, such as phytosterols and flavonoids. These nutrients may possess health benefits by producing synergistic effects and/or interactions with each other. Hence this study was intended to investigate the neurochemical pathways involved in the enhancement of memory by these nutritious nuts. The study was further aimed to examine food intake, body weight and plasma cholesterol levels following long term administration of almonds in rats.
Material and methods
Animals
The Animal Care Ethical Committee of the University had approved this investigation. Locally bred male Albino Wistar rats with mean weight 200 g were used for the experiment. Animals were caged individually in plastic cages with cubes of standard rodent diet and tap water ad libitum for 3 days before starting the experiment. Body weight and food intake of all rats were monitored during the experimental period.
Experimental protocol
Shelled almonds of best quality were purchased from the local super market which were then peeled, finely crushed and refrigerated in air tight bottle. 80 mg20 finely crushed almond was prepared as suspension in deionized water which was then orally administered to the rats.
In the beginning of experiment animals were divided into two groups, control and test. Rats were given standard rodent diet and water throughout the experiment. Fresh almond suspension was prepared daily. Body weight and food intake of all rats were monitored. Control rats (n = 6) were given deionized water and test rats (n = 6) were given almond suspension orally with the help of feeding tube. This treatment was continued for 28 days.
Food intake and body weight were monitored till almond administration. After 28 days of administration, memory test was performed and rats were decapitated. Brain and plasma samples of both groups were stored at -70o C. Tests were conducted in a noise-free environment between 9:00-13:00 hrs. Memory assessment by RAM was carried out after performing EPM test.
Memory function test by Elevated Plus Maze (EPM)
EPM was used as a behavioral model to evaluate memory in rats. The apparatus constructed of Perspex plastic with 4 arms of 50 x 10 cm area. The two enclosed arms had side walls of 40 cm high. The open and closed arms were connected with a central square (5 x 5 cm) to give the apparatus a plus sign appearance. The whole maze was raised 60 cm above the floor. The maze was placed in the same position throughout the test in laboratory where extra maze cues were there to facilitate learning. The procedure and technique was same as reported earlier by Haider et al.18 Briefly, the test comprised of three days, first day was training session while the next two days were considered as test sessions. In the training session each rat was placed in the central square and allowed to explore the EPM for 10 min and then returned to the home cages. During test sessions cut off time was 5 min and time spent in open arm was recorded. A significant decrease in time spent in open arm on subsequent EPM exposure was taken as an index of successful memory retention. This is based on the idea that during repeating testing on EPM rat acquires information about the spatial environment and avoids the elevated and open arms of the maze and prefers to stay in the closed arms where it could be safe on the maze. Total time spent in the open arm measured on the first day served as an index of learning and acquisition, whereas on the 2nd day it served as an index of retention of learned task (memory) and on the 3rd day further served as the index of consolidation of memory. Memory was measured by the degree to which the rat remembers and avoids the elevated and unenclosed arms of the maze and prefers to stay in the closed arms.
Memory function test by Radial Arm Maze (RAM)
Spatial working memory testing using eight arm radial maze task was also conducted as described by Neese et al.21 with slight modifications.22 The maze utilized in this research study consisted of a central platform, 32 cm in diameter, with 8 arms of equal length (58 cm) and width (12 cm) extending radially. Each arm had side walls of 38 cm high. The apparatus was mounted on a table so that it was 58 cm off the ground. Rats were freely allowed to visit all arms to get the food reward, which was placed near the end of one of the arm in a small plastic receptacle, this was habituation phase for 20 min. After 24 hrs, training was performed, during this phase all arms were blocked except one which was baited with food at small intervals in order to attract the rat to the end of the arm. Once the rat reached the end of the arm, the rat was returned to the central platform and the arm was rebaited in order to continue the training procedure. The training was continued for 15 min. At the end of 15 min food rewards were only placed at the end of the arm to train the rat to run to end of the arm to receive the reward. The test session was performed after 24 hrs of training, during which the same arm was again baited with food and rest of the arms were also unblocked, rats had free access to all arms. The total cut off time was 5 min and the time elapsed before the rat entered the baited arm was recorded. Throughout the experiment, care had to be taken that the relative position of maze with respect to any object serving as visual cues in laboratory was not changed. To motivate the performance, rats were food-restricted to 50% for 1 week before subjecting them to the RAM test but had free access to water.
Plasma cholesterol estimation
Plasma cholesterol was estimated by standard laboratory method.
HPLC-EC analysis
At the end of the experiment animals were decapitated using guillotine. Brain was removed immediately and stored at -70oC for the determination of TRP, 5-HT and 5-HIAA by HPLC-EC as described earlier.23 A 5-II Shim-Pack octadecylsilane separation column of 4.0 mm internal diameter and 150mm length was used. Separation was achieved by a mobile phase containing methanol (14%), octyl sodium sulfate (0.023%) and ethylenediaminetetraaceticacid (0.0035%) in 0.1 M phosphate buffer at pH 2.9, on Schimadzu LEC 6A detector at an operating potential of 0.8 volts for biogenic amines and 1.0 volts for TRP.
Statistical analysis
The behavioral and neurochemical data were analyzed by Student's t-test. P values < 0.05 were considered significant.
Results
Food intake and body weight
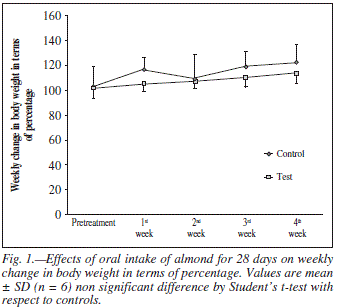
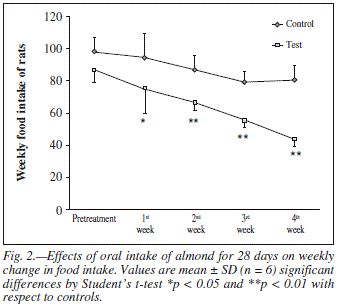
Weekly growth rate and food intake of rats following the administration of almond for 4 weeks are shown in figures 1 and 2 respectively. Test groups exhibited no differences in growth rate during the course of the study. However, there were differences in food intake, with the test group eating significantly less than the control group. Caloric intake data during the 4 weeks is summarized in table I.
Memory testing
The memory function for control and test groups following long term intake of almond measured by EPM (fig. 3) as time spent in open arm in terms of percentage. Data analyzed by Student's i-test did not reveal significant difference on the first day, however, there was significant decrease in % time spent in open arm on subsequent exposure on 2nd day (P < 0.01) and 3rd day (P < 0.01) following almond administration in rats. This observed fact taken as successful memory retention, since repeated testing on the EPM provides an index of acquisition and retention as the rat learned where it could be safe on the maze.

Spatial working memory was also observed during RAM testing following almond administration for 4 weeks (fig. 4). When data was analyzed by Student's t-test, a significant increase (P < 0.01) in memory function was observed as the time elapsed before the rat entered into baited arm was revealed significantly lower in test group than the control group.

Brain tryptophan and neurochemical estimations
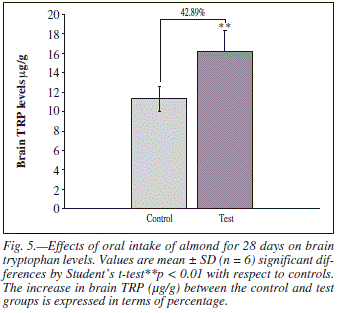
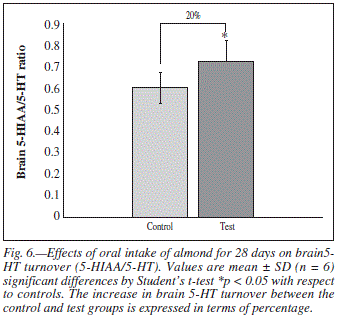
Total brain TRP (μg/g of whole brain) and 5-HT turn over (5-HIAA/5-HT ratio) were measured following 4 weeks administration of almond. The levels of TRP (P < 0.01, fig. 5) and 5-HT turn over (P < 0.05, fig. 6) both were increased significantly in the whole brain after long term intake of almond. This shows that intake of almond increased the overall brain 5-HT metabolism.
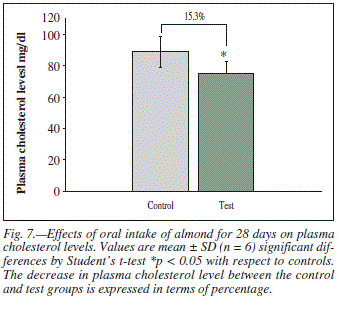
Plasma cholesterol estimation
Plasma cholesterol levels for both groups were also estimated after decapitation and shown in figure 7. Student's t-test revealed significant reduction (P < 0.05) in plasma cholesterol in test group by ~ 15% when compared with the control group.
Discussion
The finding of primary interest in the present experiment is that the long term administration of almond produces enhancing effect on memory function. This improved memory retention was evident from the gradual and significant decrease in time spent in open arm on subsequent exposures to EPM (fig. 3). Results of RAM also emphasized the same assumption as there was significant decrease in time elapsed before the rat entered the baited arm (fig. 4). Long term intake of almond was also showed to significantly increase whole brain serotonin metabolism. This can be explainable in terms of increased brain TRP concentration and 5-HT turnover in almond treated rats (fig. 5 and 6).
Memory function in rats was assessed by EPM and RAM. Both paradigms are widely used for the screening of nootropic substances.20,22,24,25 The present study showed that almond has an augmenting effect on memory retention in rats. The results are in agreement with the previously reported studies. It has been demonstrated that administration of almond improved scopolamine induced amnesia in rats.26 Central serotonin is considered as one of the neurotransmitter involved in regulation of cognitive functions. A decrease in the serotonin is believed to be involved in the occurrence of amnesia in impaired cognitive diseases.27,28 On the other hand it has been reported that increased brain 5-HT improves cognitive performance.7,29 In the present work almond administration for 4 weeks increased brain 5-HT turnover. This increased turnover was associated with an increase in the level of its precursor, TRP. The 5-HT biosynthetic enzyme, TRP hydroxylase exists in unsaturated state with its substrate. Therefore, the rate of 5-HT synthesis depends upon the brain concentration of its precursor. Increased 5-HT turnover are often taken as a measure of increased 5-HIAA levels and ultimately increased 5-HT release. An increased brain 5-HT turnover exhibited by almond-treated rats indicates an increased release of 5-HT in these rats and hence greater availability of 5-HT towards its respective receptors. Almond is rich in TRP, which is an essential amino acid, because it cannot be synthesized in vivo and must be obtained from diet. It has been shown that increased dietary availability of TRP increases 5-HT metabolism and enhances cognitive performance.9 Therefore, in the current study long term administration of almond increased brain 5-HT metabolism by elevating TRP concentration and 5-HT turnover in the brain and ultimately improved memory in rats.
The plausible active constituents of almond are polyunsaturated fatty acids (PUFAs) and choline that could be involved in memory enhancement in the present study. Dietary PUFAs [a linolenic acid; 18:3 (n-3) and linoleic acid; 12:2 (n-6)] are the main essential fatty acids that serve as precursor for fatty acids component of neuronal membrane. These fatty acids are important for the structural integrity of cell membranes and for neurotransmission, signaling and modulation of enzymatic activity.31 The PUFAs have shown to be involved in serotonin neurotransmission32 and in increased memory function.33 In addition to serotonin, acetylcholine is also considered as an important neurotransmitter involved in the regulation of cognitive function.34 Studies have revealed that consumption of essential fatty acids can effectively modulate cholinergic system.35 Acetylcholine is synthesized from substrates choline and acetyl CoA by the enzymatic action of choline acetyltransferase. Studies have indicated that increased dietary choline supplementation increases the synthesis of acetylcholine and facilitates memory function. Cohen and Wurtman36 showed that dietary intake of choline at increasing doses elevated serum choline, brain choline and brain acetylcholine concentrations in rats. Recently, a study examined the effect of dietary choline intake on memory in nondemented individuals and it was found that individuals with higher choline intake performed better on the memory tests.37 Although we did not estimate the choline and acetylcholine concentrations in the present study but from these evidences it can be suggested that in addition to TRP, important biological constituents of almond such as PUFAs and choline may also be involved in memory improving effects of almond following its long term administration in rats.
Since almonds are high in fats (55%)13 and energy dense food therefore consumers regard them as fattening foods. In the present study body weight and food intake were also measured during the course of the treatment and at the end plasma cholesterol levels were estimated in both control and test groups. After long term almond administration the body weight remained unaffected (fig. 1) while the food intake and plasma cholesterol levels were significantly decreased (table I, fig. 2 and fig. 7). The suppression in food intake in test animals may be attributed to observe increased serotonin metabolism. Several lines of evidence demonstrate that serotonergic system is recruited to reduce meal size.38,39 Serotonin modulating drugs such as 5-HT reuptake inhibitor/5-HT releaser fenfluramine has been used for the management of obesity.40 It is possible that regular intake of almonds may satisfy the appetite quickly and may decrease the food intake and ultimately decrease the plasma cholesterol level by limiting the intake of other energy-dense food.41 Hence, despite the fact that almond is rich in fats but this fatty food was not shown to exert adverse effects on body weight and plasma lipid profile16,17 as observed in the present study.
High level of cholesterol is one of the prominent risk factor for CVD. Several lines of evidence indicate that the type of fat is more important in decreasing CVD risk than the total amount of fat in the diet.42,43 According to the food composition database published by the US Department of Agriculture, 100 g of almonds contain 50.6 g of fats, majority of which are unsaturated fats.44 Unsaturated fatty acids have consistently shown to reduce total, LDL cholesterol.45 Animal studies have repeatedly performed to determine the mechanisms behind these effects and it is revealed that dietary unsaturated fatty acids enhance hepatic receptor-dependent clearance of LDL and concomitantly reduce LDL cholesterol production.46 Almonds provide a dietary source of unsaturated fatty acids which may help to reduce total cholesterol as found in the current study and may be helpful in the management of CVD.
Conclusion
In conclusion, we tested the nootropic effects of almond in male rats. The results revealed that long term almond intake enhances activity and performance in spatial memory test. Present results also indicate that almond intake increases serotonin metabolism which might be involved in memory enhancing and appetite decreasing effects. So these findings demonstrate that long term intake of almond may have beneficial effects on learning and memory processes and therefore emphasize on the use of almond as a supplement in learning and memory deficits. Almond administration also decreases food intake and total cholesterol levels. This cholesterol lowering effect may be attributed to the presence of unsaturated fatty acid content of these nuts. Therefore current findings also suggest that almond supplementation may be beneficial not only in improving memory function but also in reducing obesity and CVD risk factors.
References
1. Petersen RC, Roberts RO, Knopman DS, Boeve BF, Geda YE, Ivnik RJ, Smith GE, Jack CR Jr. Mild cognitive impairment: ten years later. Arch Neurol 2009; 66 (12): 1447-1455. [ Links ]
2. James WP. The epidemiology of obesity: the size of the problem. J Intern Med 2008; 263 (4): 336-352. [ Links ]
3. Yehuda S, Rabinovitz S, Mostofsky Dt. Essential fatty acids and the brain: from infancy to aging. Neurobiol Aging 2005; 26 (Suppl. 1): 98-102. [ Links ]
4. Spencer JP. Food for thought: the role of dietary flavonoids in enhancing human memory, learning and neuro-cognitive performance. Proc Nutr Soc 2008; 67 (2): 238-252. [ Links ]
5. Izquierdo I, Medina JH. Memory formation: the sequence of biochemical events in the hippocampus and its connection to activity in other brain structures. Neurobiol Learn Mem 1997; 68 (3): 285-316. [ Links ]
6. Schmitt JA, Wingen M, Ramaekers JG, Evers EA, Riedel WJ. Serotonin and human cognitive performance. Curr Pharm Des 2006; 12 (20): 2473-2486. [ Links ]
7. Haider S, Khaliq S, Ahmed SP, Haleem DJ. Long-term tryptophan administration enhances cognitive performance and increases 5HT metabolism in the hippocampus of female rats. Amino Acids 2006; 31 (4): 421-425. [ Links ]
8. Haider S, Khaliq S, Haleem DJ. Enhanced serotonergic neurotransmission in the hippocampus following tryptophan administration improves learning acquisition and memory consolidation in rats. Pharmacol Rep 2007; 59 (1): 53-57. [ Links ]
9. Silber BY, Schmitt JA. Effects of tryptophan loading on human cognition, mood, and sleep. Neurosci Biobehav Rev 2010; 34 (3): 387-407. [ Links ]
10. Wyatt SB, Winters KP, Dubbert PM. Overweight and obesity: prevalence, consequences, and causes of a growing public health problem. Am J Med Sci 2006; 331 (4): 166-174. [ Links ]
11. Adams KF, Schatzkin A, Harris TB, Kipnis V, Mouw T, Ballard-Barbash R, Hollenbeck A, Leitzmann MF. Overweight, obesity, and mortality in a large prospective cohort of persons 50 to 71 years old. N Engl J Med 2006; 355 (8): 763-778. [ Links ]
12. Maher VM, Brown BG, Marcovina SM, Hillger LA, Zhao XQ, Albers JJ. Effects of lowering elevated LDL cholesterol on the cardiovascular risk of lipoprotein(a). JAMA 1995; 274 (22): 1771-1774. [ Links ]
13. Davidson A. The Oxford companion to food. Oxford University press, UK 1999. [ Links ]
14. Rajaram S, Sabate J. Nuts, body weight and insulin resistance. Br J Nutr 2006; 96 (Suppl. 2): 79-86. [ Links ]
15. Garcia-Lorda P, Megias Rangil I, Salas-Salvadö J. Nut consumption, body weight and insulin resistance. Eur J Clin Nutr 2003; 57 (Suppl. 1): 8-11. [ Links ]
16. Kris-Etherton PM, Hu FB, Ros E, Sabate J. The role of tree nuts and peanuts in the prevention of coronary heart disease: multiple potential mechanisms. J Nutr 2008; 138 (9): 1746-1751. [ Links ]
17. Sabate J, Ang Y. Nuts and health outcomes: new epidemiologic evidence. Am J Clin Nutr 2009; 89 (5): 1643-1648. [ Links ]
18. Haider S, Batool Z, Tabassum S, Perveen T, Saleem S, Naqvi F, Javed H, Haleem DJ. Effects of walnuts (Juglans regia) on learning and memory functions. Plant Foods Hum Nutr 2011; 66 (4): 335-340. [ Links ]
19. Esfahlan AJ, Jamei R, Esfahlan RJ. The importance of almond (Prunus amygdalus L.) and its by-products. Food Chemistry 2010; 120 (2): 349-360. [ Links ]
20. Willis LM, Shukitt-Hale B, Cheng V, Joseph JA. Dose-dependent effects of walnuts on motor and cognitive function in aged rats. Br J Nutr 2009; 101 (8): 1140-1144. [ Links ]
21. Neese S, La GrangeL, Trujillo E, Romero D. The effect of ethanol and silymarin treatment during gestation on spatial working memory. BMC Complement Altern Med 2004; 4: 4-10. [ Links ]
22. Khaliq S, Haider S, Ahmed S, PerveenT, Haleem DJ. Relationship of brain tryptophan and serotonin in improving cognitive performance in rats. Pak J Pharm Sci 2006; 19 (1): 11-15. [ Links ]
23. Haleem DJ, Haider S. Food restriction decreases serotonin and its synthesis rate in the hypothalamus. Neuroreport 1996; 7 (6): 1153-1156. [ Links ]
24. Braida D, Ottonello F, Sala M. Eptastigmine improves eightarm radial maze performance in aged rats. Pharmacol Res 2000; 42 (4): 299-304. [ Links ]
25. Kumar A, Dogra S. Neuroprotective effect of carvedilol, an adrenergic antagonist against colchicine induced cognitive impairment and oxidative damage in rat. Pharmacol Biochem Behav 2009; 92 (1): 25-31. [ Links ]
26. Kulkarni KS, Kasture SB, Mengi SA. Efficacy study of Prunus amygdalus (almond) nuts in scopolamine-induced amnesia in rats. Indian J Pharmacol 2010; 42 (3): 168-173. [ Links ]
27. Meltzer CC, Smith G, DeKosky ST, Pollock BG, Mathis CA, Moore RY, Kupfer DJ, Reynolds CF 3rd. Serotonin in aging, late-life depression, and Alzheimer's disease: the emerging role of functional imaging. Neuropsychopharmacology 1998; 18 (6): 407-430. [ Links ]
28. Buhot MC, Martin S, Segu L. Role of serotonin in memory impairment. Ann Med 2000; 32 (3): 210-221. [ Links ]
29. Olivier JD, Jans LA, Blokland A, Broers NJ, Homberg JR, Ellenbroek BA, Cools AR. Serotonin transporter deficiency in rats contributes to impaired object memory. Genes Brain Behav 2009; 8 (8): 829-834. [ Links ]
30. Youdim KA, Martin A, Joseph JA. Essential fatty acids and the brain: possible health implications. Int J Dev Neurosci 2000; 18(8-9): 383-399. [ Links ]
31. du Bois TM, Deng C, Bell W, Huang XF. Fatty acids differentially affect serotonin receptor and transporter binding in the rat brain. Neuroscience 2006; 139 (4): 1397-1403. [ Links ]
32. Wills LM, Shukitt-Hale B, Joseph JA. Modulation of cognition and behavior in aged animals: role for antioxidant- and essential fatty acid-rich plant foods. Am J Clin Nutr 2009; 89 (5): 1602-1606. [ Links ]
33. Everitt BJ, Robbins TW. Central cholinergic systems and cognition. Annu Rev Psychol 1997; 48: 649-684. [ Links ]
34. Willis LM, Shukitt-Hate B, Joseph JA. Dietary polyunsaturated fatty acids improve cholinergic transmission in the aged brian. Genes Nutr 2009; 4 (4): 309-314. [ Links ]
35. Cohen EL, Wurtman RJ. Brain acetylcholine: control by dietary choline. Science 1976; 191 (4227): 561-562. [ Links ]
36. Poly C, Massaro JM, Seshadri S, Wolf PA, Cho E, Krall E, Jacques PF, Au R. The relation of dietary choline to cognitive performance and white-matter hyperintensity in the Framingham Offspring Cohort. Am J Clin Nutr 2011; 94 (6): 1584-1591. [ Links ]
37. Lee MD, Somerville EM, Kennett GA, Dourish CT, Clifton PG. Tonic regulation of satiety by 5-HT receptors in the mouse: converging evidence from behavioural and c-fosimmunoreactivity studies? Eur J Neurosci 2004; 19 (11): 3017-3025. [ Links ]
38. Hayes MR, Covasa M. CCK and 5-HT act synergistically to suppress food intake through simultaneous activation of CCK-1 and 5-HT3 receptors. Peptides 2005; 26 (11): 2322-2330. [ Links ]
39. Miller KJ. Serotonin 5-HT2c receptor agonists: potential for the treatment of obesity. Mol Interv 2005; 5 (5): 282-291. [ Links ]
40. Kirkmeyer SV, Mattes RD. Effects of food attributes on hunger and food intake. Int J Obes Relat Metab Disord 2000; 24 (9): 1167-1175. [ Links ]
41. Hu FB, van Dam RM, Liu S. Diet and risk of Type II diabetes: the role of types of fat and carbohydrate. Diabetologia 2001; 44 (7): 805-817. [ Links ]
42. Kris-Etherton PM, Pearson TA, Wan Y, Hargrove RL, Moriarty K, Fishell V, Etherton TD. High-monounsaturated fatty acid diets lower both plasma cholesterol and triacylglycerol concentrations. Am J Clin Nutr 1999; 70 (6): 1009-1015. [ Links ]
43. Ros E and Mataix J. Fatty acid composition of nuts - implications for cardiovascular health. Br J Nutr 2006; 96 (Suppl. 2):29-35. [ Links ]
44. Wahrburg U. What are the health effects of fat? Eur J Nutr 2004; 43 (Suppl. 1): 6-11. [ Links ]
45. Woollett LA, Spady DK, Dietschy JM. Saturated and unsaturated fatty acids independently regulate low density lipoprotein receptor activity and production rate. J Lipid Res 1992; 33 (1):77-88. [ Links ]
![]() Correspondence:
Correspondence:
Saida Haider
Neurochemistry and Biochemical Neuropharmacology Research Unit
Department of Biochemistry
University of Karachi
75270 Karachi. Pakistán
E-mail: saida-h1@hotmail.com
Recibido: 29-VIII-2012
Aceptado: 11-IX-2012

