










My SciELO
 Custom services
Custom servicesServices on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkNutrición Hospitalaria
On-line version ISSN 1699-5198Print version ISSN 0212-1611
Nutr. Hosp. vol.28 n.5 Madrid Sep./Oct. 2013
https://dx.doi.org/10.3305/nh.2013.28.5.6753
ORIGINAL / Alimentos funcionales
Nori- and Sea Spaghetti- but not Wakame-restructured pork decrease the hypercholesterolemic and liver proapototic short-term effects of high-dietary cholesterol consumption
El consumo de carne de cerdo reestructurada conteniendo Nori o, Espaguetti de Mar, pero no el de Wakame, reduce los efectos hipercolesterolemiantes y proapoptóticos hepáticos inducidos por altas cantidades de colesterol en la dieta
Adriana R. Schultz Moreira1, Juana Benedi2, Sara Bastida1, Isabel Sánchez-Reus3 and Francisco J. Sánchez-Muniz1
1Departamento de Nutrición y Bromatología I (Nutrición). Facultad de Farmacia. Universidad Complutense de Madrid. Madrid. Spain
2Departamento de Farmacología. Facultad de Farmacia. Universidad Complutense de Madrid. Madrid. Spain
3Departamento de Bioquímica y Biología Molecular II. Facultad de Farmacia. Universidad Complutense de Madrid. Madrid. Spain
This study was supported by the Spanish project AGL-2011-29644-C02-02 and Consolider-Ingenio 2010 # CSD2007-00016. We gratefully acknowledge the predoctoral fellowship awarded to Adriana Schultz Moreira by the Universidad Complutense, Madrid, Spain.
ABSTRACT
Restructured pork (RP) enriched in Seaweeds are potential functional foods. The antiapoptotic and hypocholesterolemic effects of consuming cholesterol enriched diets containing Wakame-RP (CW), Nori-RP (CN) and Sea Spaghetti (CS) were tested in a 1-wk study. Groups of six rats per group were fed a mix of 85% AIN-93M rodent-diet containing cholesterol and cholic acid as a cholesterol rising agent plus 15% RP containing alga. These diets were compared to control-RP diets enriched or not in cholesterol (CC and C, respectively). After 1-wk, cholesterol feeding significantly increased liver apoptosis markers which were significantly reduced by CS (cellular cycle DNA, caspase-3, and cytochrome c), CN (caspase-3 and cytochrome c) and CW (caspase-3) diets. CN and CS diets significantly blocked the cholesterolaemic rising effect observed in the CC group but no protective effect was observed in the CW group. Differences in seaweed composition added to RP appear responsible for blocking or not the proapoptotic and hypercholesterolemic effects of high cholesterol-RP consumption; thus, any generalization on seaweed effects or food containing seaweeds must be avoided. Although present results are worthy, future studies are demanded to ascertain the utility of consuming algal-RP as part of usual diets.
Key words: Alga. Apoptosis. Cholesterol. Functional foods. Liver damage. Restructured pork.
RESUMEN
La incorporación de algas, ricas en fibra y compuestos asociados, a reestructurados de carne de cerdo (R) resulta en derivados cárnicos potencialmente funcionales. En este trabajo se estudian los efectos antiapoptóticos e hipocolesterolemiantes de dietas en las que se incluyen agentes hipercolesterolemiantes y R enriquecidos en diferentes algas, Wakame (RW), Nori-R (RN) y Espagueti de Mar (RE). Durante una semana grupos de seis ratas cada uno recibieron una mezcla constituida por 85% de dieta AIN-93M para roedores suplementada con colesterol y ácido cólico, como agente hipercolesterolemiante, y 15% de RW, RN o RE. Estas dietas fueron comparadas con otras a las que se incorporó R control y a las que se añadió o no el agente hipercolesterolemiante. Después de 1 semana de tratamiento el incremento de marcadores de apoptosis hepática observado en el lote control con colesterol se redujo por el consumo de las dietas RE (DNA ciclo celular, caspasa-3, y citocromo c), RN (caspasa-3, y citocromo c) and RW (caspasa-3). Sólo las dietas con RN and RE bloquearon significativamente la inducción hipercolesterolemiante de la dieta control enriquecida en colesterol. Teniendo en cuenta las diferencias observadas entre los lotes respecto a sus efectos hipocolesterolémicos y antiapoptóticos, debe evitarse cualquier generalización sobre el consumo de algas y en particular de carnes conteniendo algas. Aunque los resultados son relevantes, deben realizarse estudios futuros para determinar la utilidad del consumo de estos R enriquecidos en algas dentro de dietas habituales.
Palabras clave: Algas. Alimentos funcionales. Apoptosis. Carne de cerdo reestructurada. Colesterol. Daño hepático.
Abbreviations
C: Control-group.
CC: Cholesterol-control.
CN: Cholesterol-Nori.
CS: Cholesterol-Sea spaghetti.
CW: Cholesterol-Wakame.
PUFA: Polyunsaturated fatty acids.
RP: Restructured pork.
Introduction
Seaweeds have gained importance as healthy foodstuffs in Western countries.1 High consumption of red meats has been related to increased incidence and prevalence of degenerative diseases.23 Pork-based functional foods containing seaweeds is an opportunity to improve the "image" of the meat product as these marine vegetables would positively modify meat composition increasing omega-3 fatty acid,4 fibre and associate bioactive ingredient contents5,6 and, therefore, exert positive effects on cardiovascular disease risk factors.7,8
Cholesterol consumption has been found to produce hepatomegalia, liver-fat infiltration and to increase some liver damage markers.9,10 The benefits of seaweed consumption on liver disease have been suggested; Ecklonia cava11 and Spirulina12 were found to be effective in preventing the development of hepatic fibrosis. However, no rat-liver damage protection was observed after 5-wk on diets based on restructured pork (RP) containing seaweed and highly enriched in cholesterol.10In addition, previous results13-18 suggest that generalization on the effects of seaweeds is misleading as these vegetables display differential effects attributable to differences in their composition (e.g. total and soluble dietary fibre, mineral, vitamin, and phytochemical contents).
On the other hand, the liver is the main detoxifying organ for many toxic substances and drugs that contribute to oxidative stress.19 Very recently, our group has reported that the hypocholesterolemic effect of Seaweed-RP would have a double-edge effect probably related to the CYP P450 hemo enzyme prooxidant effect.10 Indications of adverse effects associated with herbal medications, which include among others, liver failure, toxic hepatitis, and death have been reported.20Increased intake of polyunsaturated fatty acids (PUFAs) induces fatty acid oxidation in the liver and increase lipid peroxidation and cause severe hepatic injury.21Hepatocellular injury is accompanied by activation of the apoptotic pathways.22 Cellular cycle modifications and changes in caspase-3 as indicators of apoptosis level have been used to demonstrate cellular and tissue injury.23,24 To the best of our knowledge no studies on these liver damage markers have been performed in rats fed diets containing algal-RP, whether or not they were enriched in cholesterol.
As some seaweeds are hypocholesterolaemic and also antioxidants,18,25 and the cholesterol elimination via cholic acid and bile induces prooxidant stress,10 the hypothesis of the present paper is that, in the short-term, seaweed-enriched-RP check liver-cell cycle modification and damage induced by cholesterol consumption. Moreover, the protective effects are different and depend on the seaweed included in the RP. Thus, the objective of this paper was to study the short-term protective effect on the liver cellular cycle and apoptosis (measured as liver cytochrome c expression and caspase-3 activity) in Wistar rats which consumed algal-RP as part of hypercholesterolaemic diets.
Materials and methods
Materials
Wakame (Undaria pinnatifida), Nori (Porphyra umbilicalis) and Sea Spaghetti (Himanthalia elongata) were obtained from a local supplier (Algamar C.B., Redondela, Pontevedra, Spain). These seaweeds were ground in a mill (ZM 200, Retsch GmbH and Co., KG, Haan, Germany), passed through a screen with an aperture of 0.25 mm and stored in plastic flasks at 4 ± 2o C until used. Raw meat materials (post-rigor pork and pork backfat), seaweeds and additives (sodium chloride, sodium tripolyphosphate and sodium nitrite) were used as reported by Cofrades et al.26 Table I gives information on protein, fat, soluble and insoluble fibre, and ash of the different RP employed for diet preparation.
Diet preparation and experimental design
Eight experimental semi-synthetic diets (table I) were prepared: (a) the control RP diet (C) contained 85% rodent feed (AIN-93 M, purified rodent diet; Dyets #180729, DYETS, Inc., Bethlehem, PA, USA) and 15% freeze-dried control RP to which 4% cellulose had been added; (b) the Cholesterol-control RP diet (CC) was identical to the C diet but with 2.43% cholesterol (95%-98% purity) and 0.49% cholic acid (> 98% purity) substituting an equal amount of starch (AIN-93 M # 180730 diet); (c) the cholesterol Wakame RP diet (CW) consisted of a mixture of AIN-93 M #180730 feed (85%) and freeze-dried Wakame RP (15%); (d) the Cholesterol-Nori RP diet (CN) was a mixture of AIN-93 M #180730 feed (85%) and freeze-dried Nori RP (15%); (d) the cholesterol-Sea Spaghetti RP diet (CS) consisted of a mixture of AIN-93 M #180730 feed (85%) and freeze-dried Sea Spaghetti RP (15%). Details of experimental diet composition are summarized in table I.
This experiment was approved by the Spanish Science and Technology Advisory Committee and by an ethics committee of the Universidad Complutense de Madrid (Spain). All experiments were performed in compliance with Directive 86/609/EEC of November 24, 1986.
Animals and maintenance
Thirty Wistar rats with a body weight of approximately 90 g at the outset, obtained from Harlan Laboratories Models, SL, Barcelona (Spain), were kept in a temperature-controlled room (22.3 ± 1.8o C) with a 12 h light-dark cycle for 1 week to adapt to environmental conditions and fed commercial rat pellets (Panlab, Barcelona, Spain). Rats were later distributed into 5 groups of 6 animals each. The Control group received the C diet while the CC, CW, CN, and CS groups received their respective experimental diets enriched in cholesterol and the different RP. Water and food were provided ad libitum. Diets contained approximately 20.7% protein, 8.7% fat, and 4.2% total dietary fibre. At the end of the experiment, rats were anaesthetised with an intraperitoneal injection of sodium pentobarbital (45 mg/kg body weight) and euthanized by blood extraction from descendent aorta.
Food consumption and body weight change
Food intake was checked daily and body weights were measured at the start and end of the experimental week.
Plasma cholesterol
Plasma cholesterol was determined by the enzymatic colorimetric method of SPINREACT (Sant Esteve de Bas, Girona, Spain).
Liver perfusion
Livers were prepared for perfusion, caspase-3 activity, and flow cytometry analysis of DNA content, taking one animal at a time from each of the 5 experimental groups. Hepatocytes were isolated from rats by the collagenase perfusion technique as previously described.28 The viability of isolated hepatocytes (> 90%) was assessed by trypan blue exclusion as previously described.29
DNA content and cellular cycle evaluation
106 isolated viable hepatocytes were stained with propidium iodide following the multistep procedure of Vindelov et al.30 The emitted fluorescence of the DNA-propidium iodide complex was assayed in a FACScan flow cytometer (Becton-Dickinson, San Jose, California, USA) in the FL2-A channel. A double discriminator module was used to distinguish between signals coming from a single nucleus and those produced by nuclear aggregation. Data analysis was carried out by evaluation of single inputs (104 nuclei/assay) and was expressed as the percentage of DNA distribution in G0/G1 diploid population (2C), S1 population (2C - 4C), G2 + M tetraploid population (4C) phases of the cell cycle. The C DNA index was calculated as G0/G1 diploid population mean seaweed-RP divided by G0/G1 diploid population mean of C-RP. CC DNA index was calculated as G0/G1 diploid population mean seaweed-RP divided by G0/G1 diploid population mean of CC-RP.
Liver Caspase-3 activity assay
The caspase-3 activity was measured as a marker of cell apoptosis. Fluorometric assays of caspase-3 activity were conducted as described previously,31 with some modifications, in 96-well clear-bottom plates, and all measurements were carried out in triplicate wells. To each well, 100 μl of assay buffer (50 mM HEPES, pH 7.4, 10% glycerol, 10 mM dithiothreitol) was added. The peptide substrate for caspase-3 (AC-DEVD-AMC) was added to each well up to a final concentration of 20 μM. Cells were lysed with 100 μl of lysis buffer, vortexed for 30 min and centrifuged at 13,000 g for 5 min at 4o C. Protein concentrations in the supernatants were determined using the Bradford method.32 Cell lysates (20 μg protein) were added to start the reaction. Fluorescence was measured on a fluorescence plate reader at 360 nm excitation and 460 nm emission after 1 hour in incubation at 37o C. Caspase activity was calculated as [(mean AMC fluorescence from triplicate wells)-(background fluorescence)]/μg protein.
Assesment of liver apoptosis of cytochome c
Tissue samples were homogenized for 5 min in PBS buffer at a ratio of 100 mg tissue/0.5 mL PBS. The homogenates were placed on ice and then centrifuged at 12,000 g for 30 min. 40 μg of protein sample was loaded and analysed by Western blot analysis. Briefly, proteins were separated by 12% SDS-PAGE and transferred onto PVDF membranes (Millipore, Belford, NJ, USA). Membranes were blocked with blocking buffer (PBS-T containing 0.1% Tween 20) for at least 1 h with primary antibody in the above solution on an orbit shaker at 4o C overnight. Incubation was followed by washing with PBS and incubation with horseradish peroxidase-conjugated secondary antibody (anti-rabbit) in 1:1000 dilution for 1 h at room temperature. β-actin antibody (Santa Cruz Biotechnology Inc, Santa Cruz, California, USA) was used as loading control.
Statistical analyses
All results were expressed as the means ± SD of six animals in duplicate. Differences between groups were analysed by one-way ANOVA followed by Tukey post-hoc study. Statistical analyses were conducted using the SPSS version 15.0 statistical analysis packages. Differences were accepted as significant when p ≤ 0.05.
Results
Feed intake, body weight gain and feed conversion
No significant effects were observed in feed consumption, bodyweight increase or conversion rate (p > 0.05) (table II).
Plasma cholesterol
Cholesterol in diet increased plasma cholesterolemia in all groups (p < 0.001). However, CN and CS diets induced lower plasma cholesterolemia than C and CW groups (p < 0.05) (table I).
Hepatocyte cellular cycle and DNA content
Table III shows the percentage of cell cycle populations in relation to ploidy and DNA content. For the percentage of apoptotic cells (sub G1), content of DNA lesser 2n, analyzed in the cell cycle was affected by diet (p = 0.030) and increased 2.5 fold in the CC vs. C animals. The sub G1 was significantly decreased by CS when compared to CC rats, while no difference was observed when compared to the C group. By contrast, CN and CW resulted in a decreased sub G1 although no significant respect CC groups. Non-significant differences were observed in the pattern of DNA distribution and DNA indices between groups CC DNA index.
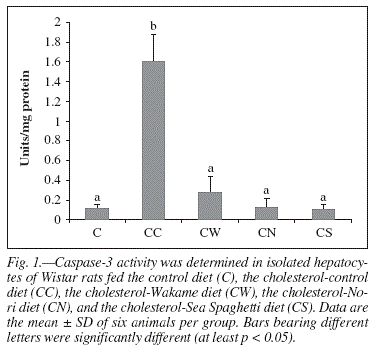
Caspase-3 activity assay
Caspase-3 activity was assayed in liver cytosolic fraction in all experimental groups and is presented in figure 1. There was a significant increase in the caspase-3 activity in hypercholesterolemic rats when compared to the control group (p < 0.05). The caspase-3 activity was significantly decreased by algae-diets when compared to hypercholesterolemic rats (p < 0.001). CS, CW, and CN displayed no differences (p > 0.05) when compared to the C group.
Cytochrome c expression
The release of mitochondrial cytochrome c into cytosol is one indicator of perturbation of mitochondrial membrane stability. We further detected cytochrome c protein levels in cytosol using western blot analysis. Immunoblot analysis showed higher release of cytosolic cytochrome c in CC rats in comparison to the C group (p < 0.05), while a consequent diminution of mitochondrial cytochrome c levels was observed when compared to the C group. Cytosolic cytochrome c protein levels analyzed by quantitative densitometry were increased by 250% (p < 0.05) in CC rats compared with the C group. The levels of cytosolic cytochrome c were decreased by CN and CS groups compared with the CC animals, while no difference was observed when compared to the C group (fig. 2).

Discussion
The experiment was designed to test possible benefits of 1-wk consumption of seaweed-RP as part of cholesterol enriched diets on liver apoptosis and DNA cell cycle. For the first time, flow cytometry analyses were tested in seaweed-RP diets both with and without added cholesterol. Rat groups present similar food acceptation and feed conversion; thus results should be related to differences in composition of seaweeds. The inclusion of cholesterol in diet promptly and highly affected the cholesterolemia; however, clear differences were observed between groups as CC increased plasma cholesterol 64%, CW 43%, and CN and CS by 25%. Semisynthetic casein-based diets with added cholesterol have been found to increase cholesterolemia in medium1,9,33 and large-term studies.16 However, the hypocholesterolaemic effects observed due to seaweed inclusion were different and highly dependent on the type of alga included in diet.1,16 According to Ren et al.,34 some seaweed polysaccharides exert hypolipemic effects in rats fed a diet rich in sodium and cholesterol. However, sodium alginate, funoran, porphyran, and carrageenan were found to interact with dietary cholesterol to facilitate its excretion, whilst dietary agar was almost inactive.35
Mammal livers contain polyploid hepatocytes, whose number depends on the animal's age and species.36 Thus, rat foetal hepatocytes are mainly diploids (85.3% of cells are in the diploid phase), 7.3% in DNA synthesis phase and 7.4% polyploids (tetraploids plus octoploids).37 In the current study, the diploid phase accounted for 73.0 to 78.6 % and tended to decrease after cholesterol feeding, while S plus tetraploid phases represented 14.1 to 19.6%. Apoptosis increased by cholesterol feeding in CC rats but CS rats showed similar apoptotic level to the C group while CW and CN intermediate values with respect to those of CC and CN groups. The necrosis-regeneration process is characterized by a change in the distribution of different populations of hepatocytes and, while normal hepatocytes have stability in their DNA content and chromosome organization, proliferative state of hepatocytes (regenerating) are involved in the replication process through the cell cycle including DNA synthesis programmed to duplicate the genomic material of the cell before its division. Regenerating cells, therefore, have a higher DNA content than quiescent cells (resting phase) and this content can vary in each cycle according to the progression of DNA synthesis. Thus, according to the cellular apoptosis/cellular DNA the inclusion of seaweed to RP diets induced moderate antiapoptotic effects, mostly in the CN diet.
Free radicals induce oxidative stress, which is balanced by the body's endogenous antioxidant systems with an input from co-factors, and by the ingestion of exogenous antioxidants. Disturbances in mitochondrial membrane permeability cause leakage of free radicals and cytochrome c from the mitochondria to the cytosol. Once cytochrome c is released into the cytosol, it binds to another protein, Apaf-1, and promotes activation of the caspase cascade, leading to cell death.38 In CC diet-fed rats, the levels of cytosolic cytochrome c and the activation of caspase-3 were much greater than those of the C group. This indicates that the release of cytochrome c into the cytosol is due to mitochondrial membrane damage, which is induced by increased levels of radicals. Cytosolic cytochrome c activates protease activators (caspases), and this may cause further apoptosis, which is also supported from the cellular cycle-DNA content. As also reported by Zamora et al.,39 dietary cholesterol increased apoptosis. However, the inclusion of alga blocked those effects in the CS group, which remained at basal levels, while CW and CN diets partially blocked the apoptotic effect of dietary cholesterol. However, seaweeds contain antioxidants and other substances that probably block the negative effect on the antioxidant status of reducing plasma cholesterol. Thus, some amino acids and antioxidant activity molecules (e.g. zinc)40 of red and brown seaweeds have been reported to be antiapoptotic.39 In addition, vitamins A and E, and some polyphenols present in seaweeds1 could act as antioxidants and have a role in the partial antiapoptotic effects observed. Fucose-containing sulphated polysaccharides extracted from Sargassum sp. and from Fucus vesiculosus respectively have been reported to induce growth inhibition and apoptosis of melanoma B16 cells in vitro.41,42
The pro-apoptotic effect of dietary cholesterol and the antiapoptotic effect of seaweed-RP diet were also supported by the results of caspase-3 activity. In fact CW, CN, and CS rats displayed much lower caspase-3 activities than their CC counterparts. Similarly, cytosolic cytochrome c expression increased 2.5-fold in CC animals and decreased to C rat values with consumption of CN and CS diets. All these results suggest that algal compounds, mostly in the case of CS and CN, blocked the proapoptotic effect of dietary cholesterol and reduced the release of mitochondrial cytochrome c into the cytosol occurring in CC rats. Thiruchenduran et al.43 reported that the increase in the expression of caspase-3 and cytochrome c observed in cholesterol fed rats was partially blocked in the grape seed proanthocyanidin-treated group via their ability to reduce, directly or indirectly, free radicals production. Although we did not measure free radical production in this short-term experiment, CN and CS diets probably reduced free radical levels via algal antioxidants and other compounds, consequently maintaining mitochondrial membrane permeability.18 This would, in turn, lead to tissue damage and/or apoptosis reduction. Mohamed et al.44reported that seaweed consumption increases the antioxidant endogenous production in vivo and that malondialdehyde were significantly reduced in liver and other organs of aging animals by porphyrans and sulphated galactans from Porphyra spp. However, a hystologycal study performed in Wistar rats fed the same RP showed, after 5-wk treatment, that the antiapoptotic and other protective effects were lost.10 As in the Schultz Moreira et al.10 study plasma cholesterol were much higher than those observed in the present study, it must be suggested that long-term cholesterol feeding must increase plasma cholesterolemia to a level that implies a high cholesterol to cholic acid conversion.44 In the cholesterol to bile acids conversion, the cytochrome P450 7A-1 hemoenzyme is induced, highly increasing the production of free radicals.45,46 This effect probably blocks the protection again apoptosis observed in the present short-term study and deserve future studies.
Conclusion
In summary, in this 1-week short-term study, sea Spaghetti-RP, and Nori-RP to a lesser extent, reduced the apoptosis induced by cholesterol feeding. More studies are needed to understand the results obtained and to ascertain the utility of consuming algal-RP as part of very cholesterol-rich diets.
References
1. Bocanegra A, Bastida S, Benedi J, Rodenas S, Sanchez-Muniz FJ. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J Med Food 2009; 2: 236-58. [ Links ]
2. Pauwels EK. The protective effect of the Mediterranean diet: focus on cancer and cardiovascular risk. Med Princ Pract 2011; 20: 103-11. [ Links ]
3. Solfrizzi V, Frisardi V, Seripa D, Logroscino G, Imbimbo BP, D'Onofrio G, Addante F, Sancarlo D, Cascavilla L, Pilotto A, Panza F. Mediterranean diet in predementia and dementia syndromes. Curr Alzheimer Res 2011; 8: 520-42. [ Links ]
4. Conchillo A, Valencia I, Puente A, Ansorena D, Astiasarán EI. Componentes funcionales en aceites de pescado y de alga. Nutr Hosp 2006; 21: 369-73. [ Links ]
5. Cofrades S, López-López I, Bravo L, Ruiz-Capillas C, Bastida S, Larrea MT, Jiménez-Colmenero F. Nutritional and antioxidant properties of different brown and red Spanish edible seaweeds. Food Sci Technol Int 2010; 5: 361-70. [ Links ]
6. Jiménez Colmenero F, Sánchez-Muniz FJ, Olmedilla-Alonso B, and collaborators. Design and development of meat-based functional foods with walnut: Technological, nutritional and health impact. Food Chem 2010; 123: 959-67. [ Links ]
7. Sánchez-Muniz FJ. Dietary fibre and cardiovascular health. Nutr Hosp 2012; 27: 31-45. [ Links ]
8. Babio N, Balanzá R, Basulto J, Bulló M, Salas-Salvadó J. Fibra dietética: influencia sobre el peso corporal, el control glicémico y el perfil del colesterol plasmático. Nutr Hosp 25: 327-40. [ Links ]
9. Sánchez-Muniz FJ, Cava F, Viejo JM, Bastida S, Higon E, Marcos A. Olive oil-fried sardines in the prevention of dietary hypercholesterolemia in rats. Effects on some serum lipids and cell-damage marker enzymes. Nutr Res 1996; 16: 111-21. [ Links ]
10. Schultz Moreira AR, García-Fernández RA, Bocanegra A, Méndez MT, Bastida S, Benedí J, Sánchez-Reus MI, Sánchez-Muniz FJ. Effects of seaweed-restructured pork diets enriched or not with cholesterol on rat cholesterolemia and liver damage. Food Chem Toxicol 2013; 56: 223-30. [ Links ]
11. Yokogawa K, Matsui-Yuasa I, Tamura A, Terada M, Kojima-Yuasa A. Inhibitory effects of Ecklonia cava extract on high glucose-induced hepatic stellate cell activation. Mar Drugs 2011; 9: 2793-808. [ Links ]
12. Kuriakose GC, Kurup MG. Antioxidant and antihepatotoxic effect of Spirulina laxissima against carbon tetrachloride induced hepatotoxicity in rats. Food Funct 2011; 2: 190-6. [ Links ]
13. Bocanegra A, Benedí J, Sánchez-Muniz FJ. Differential effects of konbu and nori seaweed dietary supplementation on liver glutathione status in normo- and hypercholesterolaemic growing rats. Br J Nutr 2006; 95: 696-702. [ Links ]
14. Bocanegra A, Nieto A, Bastida S, Benedí J, Sánchez-Muniz FJ. A Nori but not a Konbu, dietary supplement decreases the cholesterolaemia, liver fat infiltration and mineral bioavailability in hypercholesterolaemic growing Wistar rats. Br J Nutr 2008; 99: 272-80. [ Links ]
15. González-Torres L, Churruca I, Schultz Moreira AR, Bastida S, Benedí J, Portillo MP, Sánchez-Muniz FJ. Effects of restructured pork containing Himanthalia elongata on adipose tissue lipogenic and lipolytic enzyme expression of normo- and hypercholesterolemic rats. J Nutrigenet Nutrigenomics 2012; 5: 158-67. [ Links ]
16. Olivero-David R, Schultz-Moreira A, Vázquez-Velasco M, González-Torres L, Bastida S, Benedí J, Sanchez-Reus MI, González-Muñoz MJ, Sánchez-Muniz FJ. Effects of Nori- and Wakame-enriched meats with or without supplementary cholesterol on arylesterase activity, lipaemia and lipoproteinaemia in growing Wistar rats. Br J Nutr 2011; 106: 1476-86. [ Links ]
17. Miurcová L, Mach L, Orsavová J. Advances in food and nutrition research: Marine medicinal foods: Implications and applications, macro and microalgae. In: Se-Kwon, K. (Eds.), Seaweed minerals as nutraceuticals. Marine bioprocess research center, Department of Chemistry, Pukyong National University, pp. 371-86, Busan, 2011. [ Links ]
18. Moreira AS, González-Torres L, Olivero-David R, Bastida S, Benedi J, Sánchez-Muniz FJ. Wakame and Nori in restructured meats included in cholesterol-enriched diets affect the antioxidant enzyme gene expressions and activities in Wistar rats. Plant Foods Hum Nutr 2010; 65: 290-8. [ Links ]
19. Goraca A, Huk-Kolega H, Piechota A, Kleniewska PE, Skibska B. Lipoic acid - biological activity and therapeutic potential. Pharmacol Rep 2011; 63: 849-58. [ Links ]
20. Ernst E. Serious adverse effects of unconventional therapies for children and adolescents: a systematic review of recent evidence. Eur J Pediatr 2003; 162: 72-80. [ Links ]
21. Rukkumani R, Aruna K, Suresh Varma P, Padmanabhan Menon V. Hepatoprotective role of ferulic acid: a dose-dependent study. J Med Food 2004; 7: 456-61. [ Links ]
22. Malhi H, Gores GJ. Molecular mechanisms of lipotoxicity in nonalcoholic fatty liver disease. Semin Liver Dis 2008; 28: 360-369. [ Links ]
23. Bautista M, Andres D, Cascales M, Morales-González JA, Sánchez-Reus MI. Effect of gadolinium chloride on liver regeneration following thioacetamide-induced necrosis in rats. Int J Mol Sci 2010; 11: 4426-40. [ Links ]
24. Guicciardi ME, Gores GJ. Apoptosis as a mechanism for liver disease progression. Semin Liver Dis 2010; 30: 402-10. [ Links ]
25. Lordan S, Ross RP, Stanton C. Marine bioactives as functional food ingredients: potential to reduce the incidence of chronic diseases. Mar Drugs 2011; 9: 1056-100. [ Links ]
26. Cofrades S, López-López I, Solas MT, Bravo L, Jiménez-Colmenero F. Influence of different types and proportions of added edible seaweeds on characteristics of low-salt gel/emulsion meat systems. Meat Sci 2008; 79: 767-76. [ Links ]
27. López-López I, Bastida S, Ruiz-Capillas C, Bravo L, Larrea MT, Sánchez-Muniz FJ, Cofrades S, Jiménez-Colmenero F. Composition and antioxidant capacity of low-salt meat emulsion model systems containing edible seaweeds. Meat Sci 2009; 83: 492-8. [ Links ]
28. Seglen PO. Isolation of hepatocytes by collagenase perfusion. In: Tyson CA, Frazier JM, editors. Methods in toxicology in vitro biological systems. Academia Press; pp. 231-43, New York, NY, USA, 1993. [ Links ]
29. Diez-Fernandez C, Sanz N, Cascales M. Changes in glucose-6-phosphate dehydrogenase and malic enzyme gene expression in acute hepatic injury induced by thioacetamide. Biochem Pharmacol 1996; 51: 1159-63. [ Links ]
30. Vindelov LL, Christensen IJ, Nissen NI. A detergent trypsin method for the preparation of nuclei for flow cytometric. Cytometry 1983; 3: 323-7. [ Links ]
31. Bijur GN, De Sarno P, Jope RS. Glycogen synthase kinase-3beta facilitates staurosporine- and heat shock-induced apoptosis. Protection by lithium. J Biol Chem 2000; 275: 7583-90. [ Links ]
32. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976; 72: 248-54. [ Links ]
33. Sánchez-Muniz FJ, García-Linares MC, García-Arias MT, Bastida S, Viejo J. Fat and protein from olive oil-fried sardines interact to normalize serum lipoproteins and reduce liver lipids in hypercholesterolaemic rats. J Nutr 2003; 133: 2302-8. [ Links ]
34. Ren D, Noda H, Amano H, Nishino Y, Nishizawa K. Study on antihypertensive and antihyperlipidemic effects of marine algae. Fish Sci 1994; 60: 83-8. [ Links ]
35. Jiménez-Escrig A, Sánchez-Muniz FJ. Dietary fibre from Edible seaweeds: chemical structure, physicochemical properties and effects on cholesterol metabolism. Nutr Res 2000; 20: 585-98. [ Links ]
36. Saeter G, Schwarze PE, Nesland JM, Jull N, Pettersen EO, Seglen PO. The polyploidizing growth pattern of normal rat liver is replaced by divisional growth in hepatocellular nodules and carcinomas. Carcinogenesis 1988; 9: 939-45. [ Links ]
37. Díez-Fernández C, Boscá L, Fernández-Simón L, Alvarez A, Cascales M. Relationship between genomic DNA ploidy and parameters of liver damage during necrosis and regeneration induced by thioacetamide. Hepatology 1993; 4: 912-8. [ Links ]
38. Tien YC, Liao JC, Chiu C, Huang TH, Huang CY, Chang WT, Peng WH. Esculetin ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. Int J Mol Sci 2011; 12: 4053-67. [ Links ]
39. Zamora JD, Otárola IC, Brenes O. La apoptosis y su relación con diversos nutrientes. Rev Chil Nutr 2005; 32: 178-90. [ Links ]
40. Rupérez P, Ahrazem O, Leal JA. Potential antioxidant capacity of sulfated polysaccharides from the edible marine brown seaweed Fucus vesiculosus. J Agric Food Chem 2002; 50: 840-5. [ Links ]
41. Ale MT, Maruyama H, Tamauchi H, Mikkelsen JD, Meyer AS. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int J Biol Macromol 2011; 49: 331-6. [ Links ]
42. Ale MT, Maruyama H, Tamauchi H, Mikkelsen JD, Meyer AS. Fucose-containing sulfated polysaccharides from brown seaweeds inhibit proliferation of melanoma cells and induce apoptosis by activation of caspase-3 in vitro. Mar Drugs 2011; 9: 2605-21. [ Links ]
43. Thiruchenduran M, Vijayan NA, Sawaminathan JK, Devaraj SN. Protective effect of grape seed proanthocyanidins against cholesterol cholic acid diet-induced hypercholesterolemia in rats. Cardiovasc Pathol 2011; 20: 361-8. [ Links ]
44. Mohamed S, Hashim SN, Rahman HA. Seweeds: A sustainable functional food for complementary and alternative therapy. Trends Food Sci Technol 2012; 23: 83-96. [ Links ]
45. Schultz Moreira AR, Benedí J, González-Torres L, Olivero-David R, Bastida S, Sánchez-Reus MI, González-Muñoz MJ, Sánchez-Muniz FJ. Effects of diet enriched with restructured meats, containing Himanthalia elongata, on hypercholesterolaemic induction, CYP7A1 expression and antioxidant enzyme activity and expression in growing rats. Food Chem 2011; 129: 1623-30. [ Links ]
46. Micheletta F, Luliano L. Free Radical attack on cholesterol: Oxysterols as markers of oxidative stress and as bioactive molecules. Immun Endoc Metab Agents Med Chem 2006; 6: 305-16. [ Links ]
47. Tabrez S, Ahmad M. Cytochrome P450 system as potential biomarker of certain toxicants: comparison between plant and animal models. Environ Monit Assess 2013; 185: 2977-87. [ Links ]
![]() Correspondence:
Correspondence:
Francisco J. Sánchez-Muniz
Departamento de Nutrición
Facultad de Farmacia. Universidad Complutense de Madrid
28040 Madrid. Spain
E-mail: frasan@ucm.es
Recibido: 31-V-2013
Aceptado: 26-VI-2013

