










My SciELO
 Custom services
Custom servicesServices on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkNutrición Hospitalaria
On-line version ISSN 1699-5198Print version ISSN 0212-1611
Nutr. Hosp. vol.33 n.5 Madrid Sep./Oct. 2016
https://dx.doi.org/10.20960/nh.573
TRABAJO ORIGINAL / Obesidad y síndrome metabólico
Association of the rs9939609 gene variant in FTO with insulin resistance, cardiovascular risk factor and serum adipokine levels in obese patients
Asociación del polimorfismo rs9939609 en el gen FTO con la resistencia a la insulina, factores de riesgo cardiovascular y niveles de adipocitoquinas en pacientes obesos
Daniel Antonio de Luis, Rocío Aller, Olatz Izaola, David Primo and E. Romero
Centro de Investigación de Endocrinología y Nutrición. Medicine School. Universidad de Valladolid. Valladolid, Spain. Department of Endocrinology and Nutrition. Hospital Clínico Universitario de Valladolid. Valladolid, Spain
ABSTRACT
Introduction: The aim of our study was to analyze the relationship of the rs9939609 FTO gene polymorphism with insulin resistance and serum adipokine levels.
Material and methods: A population of 610 patients with obesity was analyzed in a cross sectional design. Weight, blood pressure, basal glucose, c-reactive protein (CRP), insulin, insulin resistance (HOMA), lipid profile and adipocytokines (leptin, adiponectin, resistin, TNF alpha, and interleukin 6) levels were measured.
Results: Insulin (122.2 ± 101.8 pmol/L vs. 104.5 ± 61.5 pmol/L vs. 112.1 ± 74.3 pmol/L: p < 0.05) and HOMA-IR values (4.76 ± 4.4 vs. 3.71 ± 2.5 vs. 3.76 ± 3.1; p < 0.05) were higher in TT group than AT and AA groups. Triglycerides values were higher in TT group than AA group (1.42 ± 0.71 mmol/L vs. 1.39 ± 0.69 mmol/L vs. 1.23 ± 0.64 mmol/L; p < 0.05). Adiponectin levels were lower in TT genotype group than AA genotype group (35801.2 ± 35,912.3 ng/L vs. 26,718.1 ± 36,323.1 ng/L vs. 21,112.3 ± 25,623.1 ng/L p < 0.05).
Conclusion: The FTO gene polymorphism, rs9939609, was found to be associated with insulin resistance, insulin, triglyceride and adiponectin levels in obese patients with TT variant.
Key words: Adipocytokines. Cardiovascular risk factors. Insulin resistance to rs9939609 gene variant. Obesity.
RESUMEN
Introducción: el objetivo de nuestro estudio fue analizar la relación del polimorfismo rs9939609 del gen FTO con la resistencia a la insulina y los niveles de adipocitoquinas séricas.
Material y métodos: se analizó una población de 610 pacientes con obesidad en un diseño transversal. Se registraron los valores de peso, presión arterial, glucosa basal, proteína C-reactiva (PCR), insulina, resistencia a la insulina (HOMA), perfil lipídico y adipocitoquinas (leptina, adiponectina, resistina, TNF alfa y la interleucina 6).
Resultados: los niveles de insulina circulante (104,5 + /l vs. / L 112.1 ± 74,3 pmol 61,5 pmol 122,2 ± 101,8 pmol/l vs.: p < 0,05) y HOMA-IR (4,76 ± 4,4 vs. 3,71 + 2,5 vs. 3,76 ± 3,1; p < 0,05) fueron mayores en el grupo TT que en el grupo AT+ AA. Los valores de triglicéridos fueron mayores en el grupo TT que AA (1,42 ± 0,71 mmol/l vs. 1,39 + 0,69 mmol/l vs. 1,23 ± 0,64 mmol/l; p < 0,05). Los niveles de adiponectina fueron menores en el grupo con genotipo TT que en el grupo con genotipo AA (35.801,2 ± 35.912,3 ng/l vs. 26.718,1 ± 36.323,1 ng/l vs. 21.112,3 ± 25.623,1 ng/l p < 0,05).
Conclusión: el polimorfismo del gen FTO, rs9939609, está asociado con los niveles de resistencia a la insulina, insulina, triglicéridos y de adiponectina en pacientes obesos con variante TT.
Palabras clave: Adipocitoquinas. Factores de riesgo cardiovascular. Resistencia a rs9939609. Obesidad.
Introduction
Obesity, which is a risk factor for various disorders including type 2 diabetes, hypertension, cancer, and cardiovascular disease, is one of the most common disorders in clinical practice worldwide. It has been reported that the occurrence of obesity is determined by both environmental and genetic factors (1-2). Common polymorphisms of the fat mass and obesity associated gene (FTO) have been linked to obesity in some populations (3-5). The FTOgene is highly polymorphic, and several polymorphisms of the gene have been found to be associated with obesity or obesity phenotypes, such as high body mass index (BMI). One of these genetic variants (rs9939609), located within the first FTO intron, has been related to an increased risk for both obesity and type 2 diabetes mellitus (7-16). Although the association of the FTO gene with obesity is observed across many different ethnic populations (7-16), there are several controversies. Some showed an association of the FTO gene with obesity (15-16), and some failed to replicate the result (17). The reason for this discrepancy is not clear. Lack of adjustment for confounding factors, especially dietary intake, may also be the reason, since such factors seem to be important to modulate the gene susceptibility for lifestyle-related disorders such as obesity.
In the other hand, the current view of adipose tissue is that of an active secretor organ, sending out and responding to signals that modulate appetite, insulin sensitivity, energy expenditure, inflammation and immunity. Adipocytokines are proteins produced mainly by adipocytes (18). These molecules have been shown to be involved in the pathogenesis of the metabolic syndrome and cardiovascular disease (for example; adiponectin, leptin, resistin, IL6 and TNF alpha) (19-22). Association of this FTO polymorphism with adipokine levels has been evaluated in few studies (23-25). Contradictories data have been obtained with leptin, adiponectin and interleukine-6 levels (19-25) and the relationship with resistin levels has not yet been evaluated.
Our aim was to analyze the relationship of the rs9939609 FTO gene polymorphism with body weight, insulin resistance, cardiovascular risk factors and serum adipokine levels.
Subjects and methods
SUBJECTS
A population of 610 patients with obesity was analyzed in a cross sectional design. These patients were recruited in a Nutrition Clinic Unit and signed informed consent. Local ethical committee approved the protocol (CIENC Committee Valladolid). Exclusion criteria included history of cardiovascular disease or stroke during the previous 24 months, total cholesterol > 12.8 mmol/L, triglycerides > 3.3 mmol/L, blood pressure > 140/90 mmHg, fasting plasma glucose > 3.9 mg/dL, as well as the use of sulphonilurea, thiazolidinedions, insulin, dypeptidil type 4 inhibitors, exenatide, glucocorticoids, antineoplasic agents, agiotensin receptor blockers and angiotensin converting enzyme inhibitors. Smoking habit has been excluded, too. Inclusion criteria included body mass index > 30, age > 18 years and signed informed consent.
PROCEDURE
Weight, blood pressure, basal glucose, c-reactive protein (CRP), insulin, insulin resistance (HOMA-IR), total cholesterol, LDL-cholesterol, HDL-cholesterol, triglycerides blood and adipocytokines (leptin, adiponectin, resistin, TNF alpha, and interleukin 6) levels were measured. A tetrapolar bioimpedance and a prospective serial assessment of nutritional intake with 3 days written food records were realized. Genotype of FTO gene polymorphism (rs9939609) was studied.
Genotyping of rs9939609 FTO gene polymorphism
Oligonucleotide primers and probes were designed with the Beacon Designer 5.0 (Premier Biosoft International®, LA, CA). The polymerase chain reaction (PCR) was carried out with 50 ng of genomic DNA, 0.5 uL of each oligonucleotide primer (primer forward: 5'-GGCTCTTGAATGAAATAGG-3'and reverse 5'-GACTGTTACCTATTAAAACTTTAG-3' and 0.25 uL of each probes (wild probe: 5'-Fam-ATC AAG AGC ACG GTC AAG ATT GCC-BHQ-1-3') and (mutant probe: 5'-Texas red- ATC AAG AGC ACA GTC AAG ATT GCC -BHQ-1-3') in a 25 uL final volume (Termociclador iCycler IQ (Bio-Rad®), Hercules, CA). DNA was denaturated at 95 oC for 3 min; this was followed by 35 cycles of denaturation at 95 oC for 15 s, and annealing at 55ofor 45 s). The PCR were run in a 25 uL final volume containing 12.5 uL of IQTM Supermix (Bio-Rad®, Hercules, CA) with hot start Taq DNA polymerase.
BIOCHEMICAL ASSAYS
Plasma glucose levels were determined by using an automated glucose oxidase method (Glucose analyser 2, Beckman Instruments, Fullerton, California). Insulin was measured by RIA (RIA Diagnostic Corporation, Los Angeles, CA) with a sensitivity of 3.6 pmol/L (normal range 3.5-220 pmol/L) (26) and the homeostasis model assessment for insulin resistance (HOMA-IR) was calculated using these formula (insulin x glucose/22.5) (27). CRP was measured by immunoturbimetry (Roche Diagnostics GmbH, Mannheim, Germany), with a normal range of (0-28nm l/L) and analytical sensivity 2 nmol/L. Serum total cholesterol and triglyceride concentrations were determined by enzymatic colorimetric assay (Technicon Instruments, Ltd., New York, NY, USA), while HDL cholesterol was determined enzymatically in the supernatant after precipitation of other lipoproteins with dextran sulphate-magnesium. LDL cholesterol was calculated using Friedewald formula.
Resistin was measured by ELISA (Biovendor Laboratory, Inc., Brno, Czech Republic) with a sensitivity of 200 ng/L with a normal range of 4,000-12,000 ng/L (28). Leptin was measured by ELISA (Diagnostic Systems Laboratories, Inc., Texas, USA) with a sensitivity of 50 ng/L and a normal range of 10,000-100,000 ng/L (29). Interleukin 6 and TNF alpha were measured by ELISA (R&D systems, Inc., Mineapolis, USA) with a sensitivity of 0.7 ng/L and 0.5 ng/L, respectively. Normal values of IL6 was (1.12-12.5 ng/L) and TNF alpha (0.5-15.6 ng/L) (30-31). Adiponectin was measured by ELISA (R&D systems, Inc., Mineapolis, USA) with a sensitivity of 246 ng/L and a normal range of 8,650.0-21,430.0 ng/L (32).
ANTROPOMETRIC MEASUREMENTS AND DIETARY INTAKES
Body weight was measured to an accuracy of 0.1 kg and body mass index computed as body weight/(height2). Waist (narrowest diameter between xiphoid process and iliac crest) and hip (widest diameter over greater trochanters) circumferences to derive waist-to hip ratio (WHR) were measured, too. Tetrapolar body electrical bioimpedance was used to determine body composition with an accuracy of 5 g (33). An electric current of 0.8 mA and 50 kHz was produced by a calibrated signal generator (Biodynamics Model 310e, Seattle, WA, USA). Blood pressure was measured twice after a 10 minutes rest with a random zero mercury sphygmomanometer, and averaged.
Patients received prospective serial assessment of nutritional intake with 3 days written food records. All enrolled subjects received instruction to record their daily dietary intake for three days including a weekend day. Handling of the dietary data was by means of a personal computer equipped with a software (-Dietosource 2.0®), incorporating use of food scales and models to enhance portion size accuracy. Records were reviewed by a registered dietician and analyzed with the software (Dietosource 2.0®). National composition food tables were used as reference (34).
STATISTICAL ANALYSIS
Sample size was calculated to detect differences over 0,5 kg in body weight with 90% power and 5% significance (n = 600). The results were expressed as average ± standard deviation. The distribution of variables was analyzed with -Kolmogorov-Smirnov test. Patients were divided by genotype in 3 groups (TT, TA and AA) and ANOVA test was used where indicated with Bonferroni test as a post hoc test. Non-parametric variables were analyzed with the Mann-Whitney U test. Pearson test was used to analyze correlation. Qualitative variables were analyzed with the chi-square test, with Yates correction as necessary, and Fisher's test. A p-value under 0.05 was considered statistically significant.
Results
Six hundred and ten patients gave informed consent and were enrolled in the study. No dropout during the study. The mean age was 45.3 ± 11.1 years and the mean BMI 35.7 ± 6.0.Two hundred and ninety patients (48.1%) had the genotype TT, 134 (21.7%) patients had the genotype TA and 186 patients had the genotype AA (30.2%). Age was similar in all groups (46.1 ± 16.1 years in TT groupvs.44.9 ± 16.2 years in TA group vs. 43.9 ± 15.2 years in AA group). Sex distribution was similar in different genotype groups (males vs females: 23.3% vs. 76.7% in TT group, 25.4% vs 74.6% in TA group, 30.1% vs. 69.9% in AA group).
Table I shows the anthropometric variables. No differences were detected among genotype groups.

Table II shows the classic cardiovascular risk factors. Insulin and HOMA values were higher in TT group than AT and AA groups. Triglycerides values were higher in TT group than AA group.
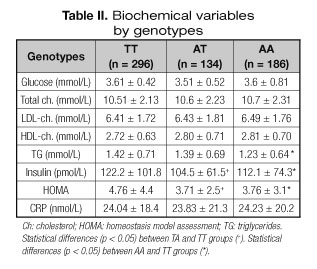
Table III shows nutritional intake with 3 days written food records. No statistical differences were detected in caloric, carbohydrate, fat, and protein intakes. Aerobic exercise per week was similar in both groups.

Table IV shows levels of adipocytokines. Adiponectin levels were lower in TT genotype group than AA genotype group. No differences were detected among genotype groups in other serum adipocytokine levels.

Discussion
We analyzed the single nucleotide polymorphism (SNP) rs9939609 of the FTO gene in obese Caucasian subjects. No associations could be found between investigated SNP and BMI, weight and some cardiovascular risk parameters (glucose, LDL cholesterol, HDL cholesterol and blood pressure). However, an association between this SNP and insulin levels, insulin resistance, triglyceride levels and adiponectin was observed.
The relation of rs9939609 with body weight is a contradictory topic area. For example, Do et al reported that the FTO variants (rs17817449 and rs1421085) have been associated with several measures of adiposity including weight, BMI, fat mass and waist circumference (35). However, rs9939609 did not show these associations (35). Dina et al. (36) note that the gene may play a role in body weight regulation, since it is highly expressed in the hypothalamic-pituitary-adrenal axis. These contradictory results might be explained by ethnicity factors. For example, Fang et al. (24) reported a significant association of FTO and BMI in Asian adults, although the risk allele effects on weight were lower in Caucasian samples, as our data shown.
In the above-mentioned study, the obesity risk alleles of rs17817449 and rs1421085 were associated with fasting insulin and HOMA-IR, the influence of these SNPs on insulin sensitivity appears to be mediated through obesity (35). We also confirmed the finding of Freathy et al (37), that FTO genotype was associated with metabolic traits. These previous results implied that the association of FTO variant with serum triglycerides and adiponectin levels may be mediated through obesity. Nevertheless, in our study, the association of these parameters was independent of body weight. In accord with other study (38), no association between leptin levels and rs9939609 genotypes were detected. In the other hand, Zimmerman et al (23) reported an association between circulating leptin levels and FTO variant, but the effect was accounted for by BMI and that the FTO A-allele tended to lower IL-6 levels. In aother pediatric group of patients (39), the minor A allele of the FTO rs9939609 was significantly associated with higher serum leptin concentrations independently of potential confounders including adiposity.
The effect of the FTO rs9939609 on insulin resistance is an unclear area, too. Tan et al (40) reported an increase in insulin resistance and hiperinsulinemia in obese patients with polycystic ovary syndrome with the A allele, without effect on glucose levels. In other study, FTO was associated with both metabolic syndrome and glucose without finding an association to insulin resistance (41). Our findings of insulin resistance and elevated triglyceride levels in patients with TT genotype were different than previous, without a clear explanation. For example, Grunnet et al. (42) have proposed that modifications of energy efficiency in oxidative muscle fibers may contribute to the association of FTO variants (A allele) and insulin resistance, but we reported an association with T allele.
These discrepancies in the metabolic findings between studies could be partly due to differences in population characteristics, such as gender, age, ethnic composition and environmental exposures such as dietary intakes. Our results suggest that there is no association of FTO with either energy intake or macronutrient composition, as other studies (43). At present the mechanism of the FTO variant on insulin resistance, triglyceride levels and adiponectin levels is uncertain. Human adipose tissue is heterogeneous in its metabolic activity, and some sites in the adipose tissue might be expanded preferentially among TT carriers, resulting in an increased insulin resistance, insulin levels and triglyceride with a decreased adiponectin levels.
Conclusion
In conclusion, the FTO gene polymorphism, rs9939609, was found to be associated with insulin resistance, insulin, triglyceride and adiponectin levels in obese patients with TT variant. A failure to control for the factors (caloric expenditure due to exercise, medications, smoking, age and gender is a biass in our design). However, further studies are necessary to confirm our results and to explore new metabolic relationships of this SNP and to performed metanalysis with pooled data as in children populations (44).
References
1. De Luis DA, González M, Aller R, Izaola O, Conde R. Effects of C358A missense polymorphism of the endocannabinoid degrading enzyme fatty acida mide hydrolase on weight loss after a hypocaloric diet. Metabolism Clinical and Experimental 2011;60;730-4. [ Links ]
2. De Luis DA, González M, Aller R, Izaola O, Conde R. Metabolic syndrome and ALA54Thr polymorphism of fatty acid-binding protein 2 in obese patients. Metabolism Clinical and Experimental 2011;60;664-8. [ Links ]
3. Hinney A, Nguyen TT, Scherag A, Friedel S, Bronner G, Muller TD, et al. Genome wide association (GWA) study for early onset extreme obesity supports the role of fat mass and obesity associated gene (FTO) variants. PLoS ONE 2007;2: e1361. [ Links ]
4. Villalobos-Comparan M, Teresa Flores-Dorantes M, Teresa Villarreal-Molina M, Rodríguez-Cruz M, García-Ulloa AC, Robles L, et al. The FTO gene is associated with adulthood obesity in the Mexican population. Obesity (Silver Spring) 2008;16:2296-301. [ Links ]
5. Song Y, You NC, Hsu YH, Howard BV, Langer RD, Manson JE, et al. FTO polymorphisms are associated with obesity but not diabetes risk in postmenopausal women. Obesity (Silver Spring) 2008;16:2472-80. [ Links ]
6. Meyre D, Delplanque J, Chèvre JC, Lecoeur C, Lobbens S, et al. Genome-wide association study for early-onset and morbid adult obesity identifies three new risk loci in European populations. Nat Genet 2009;41:157-9. [ Links ]
7. Wing MR, Ziegler J, Langefeld CD, Ng MC, Haffner SM, Norris JM, et al. Analysis of FTO gene variants with measures of obesity and glucose homeostasis in the IRAS Family Study. Hum Genet 2009;125:615-26. [ Links ]
8. Villalobos-Comparán M, Flores-Dorantes MT, Villarreal-Molina MT, Rodríguez-Cruz M, García-Ulloa AC, et al. The FTO gene is associated with adulthood obesity in the Mexican population. Obesity 2008;16:2296-2301. [ Links ]
9. Tan JT, Dorajoo R, Seielstad M, Sim XL, Ong RT, Chia KS, et al. FTO variants are associated with obesity in the Chinese and Malay populations in Singapore. Diabetes 2008:57:2851-7. [ Links ]
10. Qi L, Kang K, Zhang C, van Dam RM, Kraft P, Hunter D, Lee CH, et al. Fat mass-and obesity-associated (FTO) gene variant is associated with obesity: longitudinal analyses in two cohort studies and functional test. Diabetes 2008;57:3145-51. [ Links ]
11. Franks PW, Jablonski KA, Delahanty LM, McAteer JB, Kahn SE, Knowler WC, et al.; Diabetes Prevention Program Research Group. Diabetes Prevention Program Research Group. Assessing gene treatment interactions at the FTO and INSIG2 loci on obesity-related traits in the Diabetes Prevention Program. Diabetologia 2008;51:2214-23. [ Links ]
12. Sjögren M, Lyssenko V, Jonsson A, Berglund G, Nilsson P, Groop L, et al. The search for putative unifying genetic factors for components of the metabolic syndrome. Diabetologia 2008;51:2242-51. [ Links ]
13. Legry V, Cottel D, Ferrières J, Arveiler D, Andrieux N, Bingham A, et al. Effect of an FTO polymorphism on fat mass, obesity, and type 2 diabetes mellitus in the French MONICA Study. Metabolism 2009;58:971-5. [ Links ]
14. Yajnik CS, Janipalli CS, Bhaskar S, Kulkarni SR, Freathy RM, Prakash S, et al. FTO gene variants are strongly associated with type 2 diabetes in South Asian Indians. Diabetologia 2009;52:247-52. [ Links ]
15. Hotta K, Nakata Y, Matsuo T, Kamohara S, Kotani K, Komatsu R, et al. Variations in the FTO gene are associated with severe obesity in the Japanese. J Hum Genet 2008;53:546- 53. [ Links ]
16. Omori S, Tanaka Y, Takahashi A, Hirose H, Kashiwagi A, Kaku K, et al. Association of CDKAL1, IGF2BP2, CDKN2A/B, HHEX, SLC30A8, and KCNJ11 with susceptibility to type 2 diabetes in a Japanese population Diabetes 2008;57:791-5. [ Links ]
17. Horikoshi M, Hara K, Ito C, Shojima N, Nagai R, Ueki K, et al. Variations in the HHEX gene are associated with increased risk of type 2 diabetes in the Japanese population. Diabetologia 2007;50:2461-6. [ Links ]
18. Matsuda M, Shimomura I, Sata M. Role of adiponectin in preventing vascular stenosis. The missing link of adipo-vascular axis. J Biol CHem 2002;277:37487-91. [ Links ]
19. Kumada M, Kihara S, Sumitsuji S. Association of hypoadiponectinemia with coronary artery disease in men. Arterioscler Thromb Vasc Biol 2003;23:85-9. [ Links ]
20. Shimomoura I, Hammer RE, Ikemoto S. Leptin reverses insulin resitance and diabetes mellitus in mice with congenital lipodystrophy. Nature 1999;401:73-6. [ Links ]
21. Steppan CM, Bailey ST, Bhat S. The hormone resistin links obesity to diabetes. Nature 2001;409:307-12. [ Links ]
22. Matsuzawa Y. Adipocytokines: Emerging therapeutic targets. Current Atherosclerosis Reports 2005;7:58-62. [ Links ]
23. Zimmermann E, Skogstrand K, Hougaard DM, Astrup A, Hansen T, Pedersen O, et al. Influences of the common FTO rs9939609 variant on inflammatory markers throughout a broad range of body mass index. PLoS One 2011;6(1):e15958. [ Links ]
24. Fang H, Li Y, Du S, Hu X, Zhang Q, Liu A, et al. Variant rs9939609 in the FTO gene is associated with body mass index among Chinese children. BMC Med Genet 2010;22(11):136. [ Links ]
25. Qi L, Kang K, Zhang C, van Dam RM, Kraft P, Hunter D, et al. Fat mass-and obesity-associated (FTO) gene variant is associated with obesity: longitudinal analyses in two cohort studies and functional test. Diabetes 2008;57(11):3145-51. [ Links ]
26. Duart MJ, Arroyo CO, Moreno JL. Validation of a insulin model for the reactions in RIA. Clin Chem Lab Med 2002;40:1161-7. [ Links ]
27. Mathews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher Df. Homesotasis model assessment: insulin resistance and beta cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985;28:412-4. [ Links ]
28. Pfutzner A, Langefeld M, Kunt T, Lobig M. Evaluation of human resistin assays with serum from patients with type 2 diabetes and different degrees of insulin resistance. Clin Lab 2003;49:571-6. [ Links ]
29. Meier U, Gressner M. Endocrine regulation of energy metabolism: review of pathobiochemical and clinical chemical aspects of leptin, Ghrelin, adiponectin, and resistin. Clinical Chemstry 2004;50:1511-25. [ Links ]
30. Lubrano V, Cocci F, Battaglia D, Papa A. Usefulness of high-sensitivity IL6 measurment for clinical characterization of patients with coronary artery disease. J Clin Lab Anal 2005;19:110-4. [ Links ]
31. Khan SS, Smith MS, Reda D, Suffredini AF, Mc Coy JP. Multiplex bead array assays for detection of soluble cytokines: comparisons of sensitivity and quantitative values among kits from multiple manufactures. Cytometry B Clin Cytom 2004;61:35-9. [ Links ]
32. Suominen P. Evaluation of an enzyme immunometric assay to measure serum adiponectin concentrations. Clin Chem 2004;50:219-21. [ Links ]
33. Pichard C, Slosman D, Hirschel B, Kyle U. Bioimpedance analysis: an improved method for nutritional follow up. Clin Res 1993;41:53. [ Links ]
34. Mataix J, Mañas M. Tablas de composición de alimentos españoles. Ed: University of Granada; 2003. [ Links ]
35. Do R, Bailey S, Desbiens K, Belisle A, Montpetit A, Bouchard C, et al. Genetic variants of FTO influence adiposity, insulin sensitivity, leptin levels, and resting metabolic rate in the Quebec Family study. Diabetes 2008;57:1147-50. [ Links ]
36. Dina C, Meyre D, Gallina S. Variation in FTO contributes to childhood obesity and severe adult obesity. Nat Genet 2007;39:724-6. [ Links ]
37. Freathy RM, Timpson NJ, Lawlor DA, Pouta A. Common variation in the FTO gene alters diabetes-related metabolic traits to the extent expected given its effect on BMI: Diabetes 2008;57:1419-26. [ Links ]
38. Zabena C, González Sánchez JL, Martínez Larrad MT, Torres García A, Álvarez Fernández J. The FTO obesity gene, Genotyping and gene expression analysis in morbidly obese patients. Obes Surg 2009;19:87-95. [ Links ]
39. Labayen I, Ruiz JR, Ortega FB, Dalongeville J. Association between the FTO rs9939609 polymorphism and leptin in European adolescents: a possible link with energy balance control. The HELENA study. International Journal of Obesity 2011;35;66-71. [ Links ]
40. Tan S, Scherag A, Janssen O, Hahn S. Large effects on BMI and insulin resistance of fat mass and obesity associated gene (FTO) variants in patients with PCOS: BMC medical genetics 2010;11:12-7. [ Links ]
41. Attaoua R, Ait Sm, Radian S, Fica S, Hanzu F. FTO gene associates to metabolic syndrome un women with PCOS. Biochem Byophys Res Commun 2008;373;230-4. [ Links ]
42. Grunnet LG, Brons C, Jacobsen S, Nilsson E, Astrup A, Hansen T, et al. Increased recovery rates of phosphocreatine and inorganic phosphate after isomeric contraction in oxidative muscle fibers and elevated hepatic insulin resistance in homozygous carriers of the A-allele of FTO rs993969. J Clin Endocrinol Metab 2009;94:596-602. [ Links ]
43. Berentzen T, Kring SI, Holst C, Zimmermann E, Jess T, Hansen T, et al. Lack of association of fatness-related FTO gene variants with energy expenditure or physical activity. J Clin Endocrinol Metab 2008;93:2904-8. [ Links ]
44. Quan LL, Wang H, Tian Y, Mu X, Zhang Y, Tao K. Association of fat-mass and obesity-associated gene FTO rs9939609 polymorphism with the risk of obesity among children and adolescents: a meta-analysis. Eur Rev Med Pharmacol Sci 2015;19:614-23. [ Links ]
![]() Correspondence:
Correspondence:
Daniel Antonio de Luis.
Department of Endocrinology and Nutrition.
Institute of Endocrinology and Nutrition.
Medicine School.
Universidad de Valladolid.
Avda. Ramón y Cajal, 7.
47002 Valladolid, Spain
e-mail: dadluis@yahoo.es
Received: 17/03/2016
Accepted: 08/04/2016
