










My SciELO
 Custom services
Custom servicesServices on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkNutrición Hospitalaria
On-line version ISSN 1699-5198Print version ISSN 0212-1611
Nutr. Hosp. vol.34 n.6 Madrid Nov./Dec. 2017
https://dx.doi.org/10.20960/nh.981
Relación de la variante rs1800777 del gen CETP (proteína transportadora de ésteres de colesterol) con la masa grasa y HDL colesterol, en sujetos obesos con diabetes mellitus tipo 2
Relation of variant rs180077 of gen cholesterol ester transfer protein variant, with fat mass, HDL-cholesterol in obese subjects with diabetes mellitus type 2
Daniel A. de Luis, Olatz Izaola, David Primo, Susana García Calvo, Emilia Gómez Hoyos, Juan José López Gómez, Ana Ortola, Cristina Serrano, Esther Delgado y Beatriz Torres Torres
Centro de Investigación de Endocrinología y Nutrición. Departamento de Endocrinología e Investigación. Hospital Clínico Universitario. Facultad de Medicina. Universidad de Valladolid. Valladolid
Dirección para correspondencia
RESUMEN
Antecedentes: existe poca evidencia sobre el papel de los polimorfismos de CETP en sujetos obesos diabéticos.
Objetivos: evaluar la asociación del polimorfismo (rs1800777) del gen CETP sobre parámetros antropométricos, perfil lipídico y adipocitoquinas en pacientes obesos con diabetes mellitus.
Material y métodos: un total de 229 obesos con diabetes mellitus tipo 2 fueron reclutados. Una impedancia bioeléctrica, la presión arterial, ingesta dietética, ejercicio y bioquímica fueron analizados.
Resultados: un total de 217 pacientes (94,8%) presentaron el genotipo GG y 12 pacientes GA (5,2%) (no se detectó el genotipo AA). El peso (delta: 14,4 ± 2,1 kg, p = 0,01), índice de masa corporal (delta: 2,2 ± 1,1 kg/m2, p = 0,01), masa grasa (delta: 11,2 ± 3,1 kg, p = 0,02), circunferencia de la cintura (delta: 3,9 ± 2,0 cm, p = 0,02), índice cintura-cadera (delta: 0,04 ± 0,02 cm; p = 0,01), triglicéridos (delta: 48,6 ± 9,1 mg / dl, p = 0,03) y leptina (delta: 58,6 ± 15,9 mg/dl, p = 0,02) fueron superiores en los pacientes con el alelo A que en los no portadores del alelo A. El HDL-colesterol fue menor en los portadores de alelo A que los no portadores (delta: 5,6 ± 1,1 mg/dl, p = 0,03). Manteniéndose las diferencias en los análisis multivariantes en los niveles de HDL colesterol, masa grasa y peso.
Conclusión: nuestros resultados muestran una asociación del polimorfismo en posición +82 del gen CETP sobre los niveles de HDL colesterol, y parámetros de adiposidad en pacientes obesos con diabetes mellitus tipo 2.
Palabras clave: Adipoquinas. Proteína transportadora de ésteres de colesterol. Obesidad. Polimorfismo. Niveles lipídicos. Diabetes mellitus.
ABSTRACT
Background: There is few evidence of cholesterol ester transfer protein (CETP) in subjects with obesity and diabetes mellitus.
Objectives: We examined the association of the polymorphism (rs1800777) of CETP gene on anthropometric parameters, lipid profile and adipokines in subjects with obesity and diabetes mellitus type 2.
Material and methods: A population of 229 obese subjects with diabetes mellitus type 2 was enrolled. An electrical bioimpedance, blood pressure, dietary intake, exercise and biochemical analyses were recorded.
Results: Two hundred and seventeen subjects (94.8%) had genotype GG and 12 GA (5.2%) (genotype AA was not detected). Weight (delta: 14.4 ± 2.1 kg, p = 0.01), body mass index (delta: 2.2 ± 1.1 kg/m2, p = 0.01), fat mass (delta: 11.2 ± 3.1 kg, p = 0.02), waist circumference (delta: 3.9 ± 2.0 cm, p = 0.02), waist to hip ratio (delta: 0.04 ± 0.02 cm; p = 0.01), tryglicerides (delta: 48.6 ± 9.1 mg / dl, p = 0.03) and leptin levels (delta: 58.6 ± 15.9 mg/dl, p = 0.02) were higher in A allele carriers than non A allele carriers. Levels of HDL-cholesterol were lower in A allele carriers than non-carriers (delta: 5.6 ± 1.1 mg/dl, p = 0.03). In regression analysis, HDl cholesterol, weight and fat mass remained in the model with the SNP.
Conclusion: Our results show an association of this CETP variant at position +82 on HDL cholesterol, levels and adiposity parameters in obese subjects with diabetes mellitus type 2.
Key words: Adipokines. Cholesterol ester transfer protein. Obesity. Polymorphism. Lipid levels. Diabetes mellitus.
INTRODUCTION
En la literatura se ha demostrado una asociación inversa e independiente entre la concentración de HDL-colesterol (HDL-C) y el riesgo de enfermedad cardiovascular (1). Está demostrado ampliamente que las concentraciones de HDL colesterol eran predictivas de eventos cardiovasculares, incluso en los individuos tratados con estatinas con concentraciones de colesterol LDL < 70 g/dl (2). EL metabolismo de HDL colesterol está muy bien descrito, existiendo dos proteínas que desempeñan un papel importante en esta vía: la proteína transportadora de casete de unión a ATP A1 (ABCA1) y la proteína de transferencia de éster de colesterol (CETP). La proteína CETP participa en el metabolismo de HDL, facilitando la transferencia de ésteres de colesterol de HDL a lipoproteínas que contienen ApoB a cambio de triglicéridos que se transfieren a HDL (3). La relación que existe entre algunos polimorfismos (SNP) del gen CETP con las concentraciones de HDL colesterol ya ha sido descrita en la literatura (4-9). Se ha descrito un polimorfismo de único nucleótido en el gen CETP (rs1800777) localizado en la región codificadora de este gen, no existiendo trabajos realizados específicamente en pacientes con obesidad y diabetes mellitus, de ahí el interés de su evaluación. El alelo menor (A) de este SNP aparece con una baja frecuencia en la población general (2-7%) (10). El alelo A se ha asociado con concentraciones más bajas de colesterol HDL (10) y mayor actividad CETP (11). En otro trabajo el alelo A de este SNP se asoció con el espesor de la pared íntima media de la carótida y la calcificación de la arteria coronaria (12). Aunque como vemos en la literatura la CETP es importante en el metabolismo de HDL y se conoce que los SNP del gen CETP se asocian con las concentraciones de HDL-colesterol, esta área de investigación es de interés porque los trabajos realizados con las variantes genéticas de este gen (4-11) no han valorado la posible relación con parámetros antropométricos, resistencia a la insulina y adipocitoquinas.
En el presente trabajo se evaluó la asociación del polimorfismo (rs1800777) del gen CETP sobre los parámetros antropométricos de la obesidad, perfil lipídico y adipocitoquinas en sujetos con obesidad y diabetes mellitus tipo 2.
MATERIAL Y MÉTODOS
Sujetos
Se analizó una muestra de 229 sujetos obesos (índice de masa corporal > 30 kg/m2) con diabetes mellitus tipo 2 diagnosticados según criterios ADA con una glucemia en ayunas superior a 126 mg/dl sin realizar test de sobrecarga oral de glucosa (13), de reciente diagnóstico sin ningún tratamiento específico para su diabetes. Estos pacientes habían sido remitidos a la consulta externa de nutrición clínica desde enero a diciembre de 2013. Todos los participantes firmaron un consentimiento informado del protocolo aprobado por el Comité de Ensayos Clínicos del Hospital. Los sujetos fueron excluidos si tenían antecedentes de enfermedad cardiovascular o accidente cerebrovascular durante los últimos 24 meses, antecedentes de cáncer sometidos a tratamiento activo, pérdida de peso de más del 5% del peso corporal en los últimos 6 meses, colesterol total > 250 mg/dl, triglicéridos > 250 mg/dl, presión arterial > 140/90 mmHg, así como el uso de metformina, sulfonilurea, inhibidores del tipo dipeptidil tipo IV, tiazolidinodionas, insulina, glucocorticoides, agentes antineoplásicos, bloqueadores de los receptores de angiotensina, inhibidores de la enzima convertidora de angiotensina, estatinas y otros fármacos antidislipidémicos.
Procedimiento
El peso, la presión arterial, glucosa basal, proteína C reactiva (PCR), insulina, resistencia a la insulina (HOMA-IR), colesterol total, colesterol LDL, colesterol HDL, triglicéridos y adipocitoquinas (leptina, adiponectina, resistina) se determinaron en todos los sujetos. Todas las determinaciones analíticas se realizaron en una muestra de sangre tras ayunas de 8 horas, y se congelaron a -80 °C para realizar en el mismo momento las determinaciones.
Se realizó también una bioimpedancia y una evaluación prospectiva de la ingesta nutricional durante 3 días. Se estudió el genotipo del polimorfismo del gen receptor CETP.
Procedimientos analíticos
Los niveles de glucosa plasmática se determinaron usando un método automatizado de glucosa oxidasa (analizador de glucosa 2, Beckman Instruments. Fullerton, California). La insulina se midió por radioinmunoensayo (RIA) con una sensibilidad de 0,5 mUI/L (rango normal de 0,5 a 30 mUI/L) (14) y la evaluación del modelo de homeostasis para la sensibilidad a la insulina (HOMA-IR) se calculó utilizando estos valores (15). La PCR se midió por inmunoturbimetría (Roche Diagnostis GmbH. Mannheim, Alemania), con un rango normal de (0-7 mg/dl) y una sensibilidad analítica de 0,5 mg/dl. Las concentraciones séricas de colesterol total y de triglicéridos se determinaron por ensayo colorimétrico enzimático (Technicon Instruments, Ltd., Nueva York, N.Y., EE. UU.), mientras que el colesterol HDL se determinó enzimáticamente en el sobrenadante después de la precipitación de otras lipoproteínas con sulfato de dextrano-magnesio. El colesterol LDL se calculó utilizando la fórmula de Friedewald (16). La adiponectina se midió mediante ELISA (R & D Systems, Inc. Mineápolis, EE. UU.) con una sensibilidad de 0,246 ng/ml y un intervalo normal de 8,65-21,43 ng/mL (17). La leptina se midió mediante ELISA (Diagnostic Systems Laboratories, Inc. Texas, EE. UU.) con una sensibilidad de 0,05 ng/mL y un intervalo normal de 10-100 ng/mL (18). La resistina se midió mediante ELISA (Biovendor Laboratory, Inc. Brno, República Checa) con una sensibilidad de 0,2 ng/mL con un intervalo normal de 4-12 ng/mL (19).
Genotipado
Los primers y sondas de oligonucleótidos se diseñaron con el programa Beacon Designer 5.0 (Premier Biosoft International®. Los Ángeles, California. EE. UU.). La reacción en cadena de la polimerasa (RCP) se llevó a cabo con 50 ng de ADN genómico, 0,5\mul de cada primer (cebador adelante: 5 ‘- ACGTTGGATGCTCTTCGACATCATCAACCC - 3' y reverso 5 ‘- ACGTTGGATGAATCCTGTCTGGGCCTCTCT - 3' en un volumen final de 2 microL. El DNA fue desnaturalizado a 95 °C durante 3 min, seguido de 50 ciclos de a 95 °C durante 15 s, y a 61,3 °C durante 45 s. La reacción de la polimerasa se llevó a cabo en un volumen final de 25 uL que contenía 12,5 uL de IQTM Supermix (Bio-Rad®. Hercules, California. EE. UU.) con la Taq ADN polimerasa.
Procedimientos antropométricos y presión arterial
También se midieron la circunferencia de la cintura (diámetro más estrecho entre el proceso xifoide y la cresta iliaca) y la cadera (diámetro más ancho sobre los trocánteres mayores), con ambos parámetros se calculó la relación cintura/cadera (ICC). El peso corporal se midió con una precisión de 0,1 kg y el índice de masa corporal se calculó como peso corporal en kg/(talla en m2) (Omrom. Los Ángeles, California. EE. UU.). La bioimpedancia eléctrica del cuerpo tetrapolar se utilizó para determinar la composición corporal con una precisión de 5 g (20) (EFG, Akern, Pi, It). La presión sanguínea se midió dos veces tras un periodo de reposo de 10 minutos con un esfigmomanómetro de mercurio (Omrom, Los Ángeles, California, EE. UU.) y se promediaron los resultados.
Hábitos alimenticios
Todos los sujetos evaluados recibieron instrucciones para registrar su ingesta dietética diaria cuantitativa durante tres días incluyendo un día de fin de semana con el fin de ajustar nuestro resultado por ingestas dietéticas. El manejo de los datos dietéticos se realizó mediante un software, incorporando el uso de escalas de alimentos y modelos para mejorar la precisión de tamaño de porción (Dietsource®, Gen, Swi). Este software utilizó las tablas de composición nacional de alimentos (21). La actividad física (minutos por semana) se evaluó mediante el test internacional de actividad física (22).
Análisis estadístico
El tamaño de la muestra se calculó para detectar diferencias superiores a 4 mg/dl en colesterol HDL con un 90% de potencia y un 5% de significación (n = 200). Los resultados se expresaron como promedio ± desviación estándar. La distribución de las variables se analizó con la prueba de Kolmogorov-Smirnov.
Las variables cuantitativas con distribución normal se analizaron con una prueba t de Student de dos colas. Se utilizaron pruebas no paramétricas en variables no normales. Las variables cualitativas se analizaron con la prueba del Chi-cuadrado, con la corrección de Yates como necesario y la prueba de Fisher. En las variables que presentaron en el análisis univariante una asociación con el genotipo se realizó un análisis multivariante ajustado por edad, sexo, peso, ejercicio e ingesta. El análisis estadístico se realizó para la combinación de GA y AA como grupo y GG genotipo como segundo grupo, con un modelo dominante. Se consideró estadísticamente significativo un valor de p inferior a 0,05.
RESULTADOS
Un total de 229 pacientes obesos con diabetes mellitus tipo 2 y sin tratamiento farmacológico firmaron el consentimiento informado y fueron reclutados en el estudio. La edad media fue de 54,2 ± 9,3 años y la media de IMC 38,7 ± 5,3. Todos los sujetos presentaron un peso estable durante el periodo de 2 semanas anteriores al estudio (variación de peso corporal, 0,16 ± 0,1 kg). Un total de 217 pacientes (94,8%) presentaron el genotipo GG y 12 pacientes GA (5,2%) (no se detectó el genotipo AA). La edad fue similar en ambos genotipos (GG, 56,4 ± 9,1 años vs. GA, 52,3 ± 7,1 años: ns). La distribución por sexo fue similar en los dos genotipos (GG, 34,6% varones vs. 65,4% mujeres vs. GA, 25,0% varones vs. 75,0% mujeres: ns). La variante estaba en equilibrio de Hardy-Weinberg (p = 0,29).
La tabla I muestra la media y la desviación estándar de los parámetros antropométricos y la presión arterial. El peso (delta: 14,4 ± 2,1 kg, p = 0,01), índice de masa corporal (delta: 2,2 ± 1,1 kg/m2, p = 0,01), masa grasa (delta: 11,2 ± 3,1 kg, p = 0,02), circunferencia de la cintura (delta: 3,9 ± 2,0 cm, p = 0,02) e índice cintura-cadera (delta: 0,04 ± 0,02 cm; p = 0,01) fueron superiores en los pacientes portadores del alelo A que en los no portadores. En los análisis de regresión múltiple ajustado por edad, sexo, peso, ejercicio e ingesta, el alelo A permaneció en el modelo final beta 12,30 kg (IC95% 7,22-18,36) para peso y masa grasa beta 7,19 kg (IC95% 4,10-12,88).
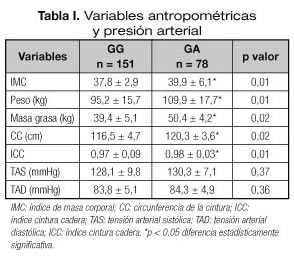
La tabla II muestra los factores clásicos de riesgo cardiovascular. Los niveles de HDL-colesterol fueron menores en los portadores de alelo A que los no portadores (delta: 5,6 + 1,1 mg/dl, p = 0,03), sin embargo los triglicéridos fueron mayores en los portadores del alelo A (delta: 48,6 + 9,1 mg/dl, p = 0,03). En los análisis de regresión múltiple ajustado por edad, sexo, peso, ejercicio e ingesta, el alelo A permaneció en el modelo final beta 4,21 mg/dl (IC95% 0,89-8,50) para HDL colesterol y no permaneció para triglicéridos beta 13,90 mg/dl (IC95% -4,19-68,13).

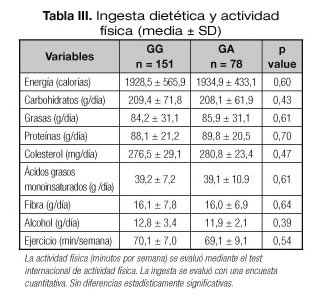
La tabla III muestra la ingesta nutricional con registros de alimentos escritos de 3 días. No se detectaron diferencias estadísticas en la ingesta de calorías, carbohidratos, grasas y proteínas. El ejercicio aeróbico por semana fue similar en ambos grupos.
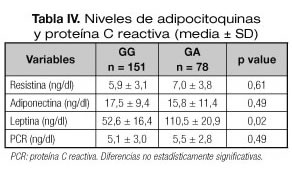
La tabla IV muestra los niveles de adipocitoquinas. Los niveles de leptina fueron superiores en los portadores del alelo A (delta: 58,6 + 15,9 mg/dl, p = 0,02). No se detectaron diferencias estadísticas en el resto de adipocitoquinas analizadas (resistina y adiponectina). En el análisis de regresión múltiple ajustado por edad, sexo, índice de masa corporal e ingesta, el alelo A no permaneció en el modelo final beta 42,1 ng/ml (IC95% -3,8-67,9).
DISCUSIÓN
El principal hallazgo de nuestro estudio fue que el SNP (rs1800777) del gen CETP se asoció con los niveles de HDL-colesterol y triglicéridos, así como con parámetros de adiposidad, como el peso, la masa grasa por impedanciometría, circunferencia de la cintura y el índice cintura cadera.
El objetivo principal de nuestro estudio transversal fue investigar la asociación del este polimorfismo con el perfil lipídico, antropometría y los factores de riesgo cardiovascular. En nuestro trabajo conseguimos demostrar una asociación inversa del alelo A en la posición +82 con los niveles de colesterol HDL y directa con los valores de triglicéridos circulantes (aunque esta asociación no se mantuvo en el análisis multivariante), así como con los parámetros antropométricos, ya que los portadores del alelo A presentaron valores más elevados de adiposidad. La frecuencia del alelo menor (A) en nuestro estudio (5,2%) es similar a la que se informó anteriormente (2-7%) (10,11). Algunos trabajos previos demostraron que algunos genotipos frecuentes de CETP reducen la actividad de CETP (23) en aproximadamente 5% a 10%, modificando los niveles del colesterol HDL entorno al 3%-5%. Un metaanálisis (23) revisó 6 genotipos de CETP incluyendo rs1800777; los resultados de este artículo sugieren que los individuos con alelos menores presentan menores niveles de colesterol HDL y un aumento del riesgo coronario. Una limitación de este metaanálisis (23) es que incluye trabajos originales que no han controlado diferentes factores del estilo de vida que influyen claramente sobre los niveles de HDL colesterol, esta falta de control podría constituir un sesgo en la asociación descrita. Los determinantes ambientales de los niveles de colesterol HDL (por ejemplo, el alcohol, el ejercicio y la ingesta de grasas en la dieta) sí que se han controlado en nuestro estudio transversal.
En un reciente estudio transversal multiétnico, Tsai y cols. (24) mostraron que el alelo A del polimorfismo rs1800777 estaba asociado con las concentraciones de HDL colesterol. En un estudio previo, el polimorfismo rs1800777 se asoció con mayor actividad de la proteína CETP y colesterol HDL más bajo, relacionándose también con estenosis carotidea y aumento de la presencia de calcio coronario (14). El mecanismo exacto por el que esta variante en la región codificadora del gen CETP influye sobre la enfermedad de la arteria coronaria permanece aún desconocido.
Hasta donde sabemos la relación entre este SNP y los marcadores de adiposidad han sido evaluados en un trabajo y no se ha encontrado una relación significativa (25); tampoco se han realizado estudios específicos en pacientes con diabetes mellitus tipo 2. No se ha evaluado la relación de este SNP con la ingesta dietética; solo hemos encontrado un trabajo en la literatura (26) en el que en un grupo de adolescentes tratados con dieta, este polimorfismo lograba explicar hasta el 2% del cambio BMI-SD después de 10 semanas de la intervención con una dieta hipocalórica. En este trabajo no se midió la circunferencia de la cintura ni la masa grasa con impedanciometría como en nuestro estudio. No obstante, la asociación con los parámetros de adiposidad, tanto al principio como después de la intervención dietética, fue estadísticamente significativa. El mecanismo molecular exacto responsable de los efectos biológicos sobre la obesidad (masa grasa, circunferencia de la cintura y relación cintura/cadera) no se ha descrito hasta la fecha. Tal vez este SNP podría influir en la diferenciación del tejido adiposo visceral, aunque otro mecanismo desconocido podría estar implicado en esta interesante relación. Por último, la relación de rs1800777 con los niveles séricos de leptina tampoco había sido descrita anteriormente. Nuestros datos podrían explicarse por la asociación del alelo menor con una mayor masa grasa en el análisis multivariante lo cual explica unos valores más elevados de leptina circulante y la pérdida de asociación con la leptina al realizar el análisis ajustado.
Nuestro trabajo sin duda presenta algunas limitaciones. En primer lugar, hay muchos factores no genéticos no controlados que podrían influir en las relaciones encontradas con nuestro diseño (estado hormonal, tipo de ejercicio, etc.). En segundo lugar, no se analizaron subtipos de partículas HDL. En tercer lugar, solo se analizó un SNP del gen CETP, por lo que otras variantes genéticas podrían estar implicadas en estas asociaciones encontradas. Finalmente, nuestro diseño es un estudio trasversal, no pudiendo inferir causalidad en los resultados obtenidos.
En conclusión, nuestros resultados muestran un papel importante de esta variante de CETP en la posición +82 sobre el colesterol HDL, la masa grasa y la obesidad central en pacientes obesos con diabetes mellitus tipo 2.
BIBLIOGRAFÍA
1. Wilson PW, Abbott RD, Castelli WP. High density lipoprotein cholesterol and mortality. The Framingham Heart Study. Arteriosclerosis 1988;8:737-41.
2. Harchaoui K, van der Steeg WA, Stroes ES, Kuivenhoven JA, Otvos JD, Wareham NJ, et al. Value of low-density lipoprotein particle number and size as predictors of coronary artery disease in apparently healthy men and women: the EPICNorfolk Prospective Population Study. J Am Coll Cardiol 2007;49:547-53.
3. Tall AR. Plasma cholesteryl ester transfer protein. J Lipid Res 1993;34:1255-74.
4. Ordovas JM, Cupples LA, Corella D, Otvos JD, Osgood D, Martinez A, et al. Association of cholesteryl ester transfer protein-TaqIB polymorphism with variations in lipoprotein subclases and coronary heart disease risk: the Framingham study. Arterioscler Thromb Vasc Biol 2000;20:1323-9.
5. Liu S, Schmitz C, Stampfer MJ, Sacks F, Hennekens CH, Lindpaintner K, et al. A prospective study of TaqIB polymorphism in the gene coding for cholesteryl ester transfer protein and risk of myocardial infarction in middle-aged men. Atherosclerosis 2002;161:469-74.
6. Fumeron F, Betoulle D, Luc G, Behague I, Ricard S, Poirier O, et al. Alcohol intake modulates the effect of a polymorphism of the cholesteryl ester transfer protein gene on plasma high density lipoprotein and the risk of myocardial infarction. J Clin Invest 1995;96:1664-71.
7. Lu H, Inazu A, Moriyama Y, Higashikata T, Kawashiri MA, Yu W, et al. Haplotype analyses of cholesteryl ester transfer protein gene promoter: a clue to an unsolved mystery of TaqIB polymorphism. J Mol Med 2003;81:246-55.
8. Eiriksdottir G, Bolla MK, Thorsson B, Sigurdsson G, Humphries SE, Gudnason V. The _629C_A polymorphism in the CETP gene does not explain the association of TaqIB polymorphism with risk and age of myocardial infarction in Icelandic men. Atherosclerosis 2001;159:187-92.
9. Agerholm-Larsen B, Tybjaerg-Hansen A, Schnohr P, Steffensen R, Nordestgaard BG. Common cholesteryl ester transfer protein mutations, decreased HDL cholesterol, and possible decreased risk of ischemic heart disease: The Copenhagen City Heart Study. Circulation 2000;102:2197-203.
10. Corbex M, Poirier O, Fumeron F, Betoulle D, Evans A, Ruidavets JB, et al. Extensive association analysis between the CETP gene and coronary heart disease phenotypes reveals several putative functional polymorphisms and gene-environment interaction. Genet Epidemiol 2000;19:64-80.
11. Agerholm-Larsen B, Tybjaerg-Hansen A, Schnohr P, Steffensen R, Nordestgaard BG. Common cholesteryl ester transfer protein mutations, decreased HDL cholesterol, and possible decreased risk of ischemic heart disease: The Copenhagen City Heart Study. Circulation 2000; 102:2197-203.
12. Kakko S, Tamminen M, Paivansalo M, Kauma H, Rantala AO, Lilja M, et al. Cholesteryl ester transfer protein gene polymorphisms are associated with carotid atherosclerosis in men. Eur J Clin Invest 2000;30:18-25.
13. Standards of Medical Care in Diabetes-2017: Summary of Revisions. Diabetes Care Volume 40, Supplement 1, January 2017.
14. Duart MJ, Arroyo CO, Moreno JL. Validation of an insulin model for the reactions in RIA. Clin Chem Lab Med 2002;40:1161-7.
15. Mathews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher Df. Homeostasis model assessment: insulin resistance and beta cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985;28:412-4.
16. Friedewald WT, Levy RJ, Fredrickson DS. Estimation of the concentration of low-density lipoprotein cholesterol in plasma without use of the preparative ultracentrifuge. Clin Chem 1972;18:499-502.
17. Suominen P. evaluation of an enzyme immunometric assay to measure serum adiponectin concentrations. Clin Chem 2004;50:219-21.
18. Meier U, Gressner M. Endocrine regulation of energy metabolism: review of pathobiochemical and clinical chemical aspects of leptin, Ghrelin, adiponectin, and resistin. Clinical Chemistry 2004;50:1511-25.
19. Pfutzner A, Langefeld M, Kunt T, Lobig M. Evaluation of human resistin assays with serum from patients with type 2 diabetes and different degrees of insulin resistance. Clin lab 2003;49:571-6.
20. Lukaski H, Johson PE. Assessment of fat-free mass using bioelectrical impedance measurements of the human body. Am J Clin Nutr 1985;41:810-7.
21. Mataix J, Mañas M. Tablas de composición de alimentos españoles. Ed: University of Granada; 2003.
22. Booth M. International Consensus Group on Physical Activity Measurement. Int J Epidemiol 1996;25(6):1312-3.
23. Thompsom A, Angelantonio E, Sarwar N, Erqou S, Saleheen D, Dulalrt R. Association of cholesteryl Ester Transfer Protein Genotypes with CETP Mass and activity, lipid levels, and coronary risk. JAMA 2008;23:2777-83.
24. Tsai M, Li N, Richey A, Shea S, Jacobs D, Tracy R, et al. Association of Genetic variants in ATP-Binding cassette A1 and cholesteryl Ester Transfer Protein and differences in lipoprotein subclasses in the Multi-Ethnic Study of Atherosclerosis. Clinical Chemistry 2009;55:481-8.
25. Moleres A, Milagro FI, Marcos A, González-Zorzano E, Campoy C, Garagorri JM, et al. Common variants in genes related to lipid and energy metabolism are associated with weight loss after an intervention in overweight/obese adolescents. Nutr Hosp 2014;30:75-83.
26. Moleres A, Milagro F, Marcos A, Gonzalez Zorzano E, Campoy C, Garagorri JM. Common variants in genes related to lipid and energy metabolism are associated with weight loss after an intervention in overweight/obese adolescents. Nutr Hosp 2014;30:75-83.
 Dirección para correspondencia:
Dirección para correspondencia:
Daniel A. de Luis.
Departamento de Endocrinología e Investigación.
Hospital Clínico Universitario.
Avda. Ramón y Cajal, 3.
47003 Valladolid
e-mail: dadluis@yahoo.es
Recibido: 30/01/2017
Aceptado: 04/06/2017

