










 Custom services
Custom services
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
In recent years, interest in the intestinal microbiota and its interactions as the host has increased 1. In particular, investigations on the role of microbiota in health regulation and onset of diseases such as inflammatory diseases, allergies, diabetes and obesity 2. It is estimated that the human intestinal microbiota is composed of 10 to 100 trillion microorganisms and possesses about 150 times more genes than the human genome 3.
With the advancement of sequencing techniques, metagenomic analyses of 16S rRNA have demonstrated a large number of bacterial genes that inhabit the human gut 4. The predominance of Firmicutes and Bacteroidetes and restricted anaerobic genera such as Bacteroides, Eubacterium, Clostridium, Ruminococcus, Peptococcus, Bifidobacterium and Fusobacterium were observed in relation to facultative anaerobes such as Lactobacillus, Escherichia, Enterobacter, Enterococcus, Klebsiella and Proteus. 4,5. However, there is no accuracy in the composition of the microbiota in humans 6, being influenced by the age, genetic and environmental factors, diet and structure of the intestinal wall of the host 7.
The intestinal microbial ecosystem has been shown essential in the performance of functions such as preservation of intestinal mucosal integrity, nutrient absorption and energy homeostasis, as well as being directly linked to the immune and nervous system 8,9. It has recently been suggested that the microbiota may play a significant role in the pathogenesis of obesity and its comorbidities 8,10,11,12,13. This is partly due to an imbalance in the homeostasis of the microbiota, known as dysbiosis, which is characterized by changes in diversity, toxin production, increased permeability, and hormonal and immunological changes, culminating in low-grade inflammatory state 14.
Lipopolysaccharide (LPS), a constituent of intestinal bacteria, can be an important inducer of the inflammatory response 15. Detected in the intestinal lumen, under normal conditions, it does not represent human health problems. However, when there is an imbalance this can be easily transferred to the circulatory system. This can lead to high plasma levels of LPS, a condition known as metabolic endotoxemia 16. In addition, there is evidence that the type of diet consumed, especially diets rich in fats, may contribute to endotoxemia 17.
Much has been discussed about the role of diet in the regulation and composition of the intestinal microbiota 14,18,19. High-fat diets are associated with a reduction in intestinal bacterial diversity 20, changes in intestinal membrane integrity, inducing increased permeability and increased LPS translocation 21, changes in the immune system and generation of systemic inflammation 22.
Therefore, the purpose of this review is to examine papers from the scientific literature in order to discuss the mechanisms and interactions between diets rich in fats, intestinal microbiota and metabolic endotoxemia.
METHODS
The research was performed in the Medline/PubMed and Scopus databases. The following terms were used in English: endotoxemia; endotoxins; lipopolysaccharides; gut microbiota and dietary fat. The literature search was conducted from January to July 2017. All articles selected and included in this article were published between 2007 and 2017. They were read and critically grouped according to their thematic and scientific relevance. From this, the sections presented in this article were created: "Dysbiosis induced by high-fat diet" and "Diet rich in lipids and metabolic endotoxemia" (Fig. 1).

HIGH-FAT DIET-INDUCED DYSBIOSIS
Well-established scientific evidence reports that excessive intake of fats and refined carbohydrates are strongly associated with obesity and metabolic diseases. However, it is recent discoveries that may also influence the composition of the intestinal microbiota of the host 23. It is known that the human intestine has trillions of microorganisms, containing more genes than the human genome itself. These microorganisms have evolved and are capable of performing specific and unique biochemical and metabolic functions to the microbial species 3.
The role of the intestinal microbiota in the human disease health process has been gaining even more focus and attention, especially on the etiology of obesity and its comorbidities 24. It is suggested that intestinal bacteria are involved in energy homeostasis and body weight control, being able to extract calories from indigestible nutrients and produce short chain fatty acids, amino acids and vitamins 25. In addition, they participate in the regulation and expression of genes that signal metabolic pathways of absorption and storage of nutrients, such as fats, in the host 26.
The interaction microorganisms and host is under a complex interface with the gastrointestinal mucosa. Its outer layer offers conditions for colonization by bacteria and may suffer direct influence of the diet 27 since bacteria that degrade mucus are influenced by high-fat diets. Thus, it is evident that diet plays an important role in the modulation of the microbial ecosystem 28.
Studies have reported that the consumption of diets with high-fat contents may alter the composition and functionality of the intestinal microbiota in mice and humans 29 30 31 32 33 34-35 (Table 1 and Table 2). Short-term dietary interventions (24 h) have been shown to rapidly change the intestinal microbiota; on the other hand, long-term dietary patterns are able to modulate the composition of the microbiota, despite the detection of resilience after induction of dietary changes 36. Factors such as age, diet time, function, and properties of the different intestinal segments may also influence microbial diversity, and when the cecum and colon mucosa are compared, the latter is more resistant to variations 25.

Table II. Review of studies investigating the dysbiosis induced by a high-fat diet in humans
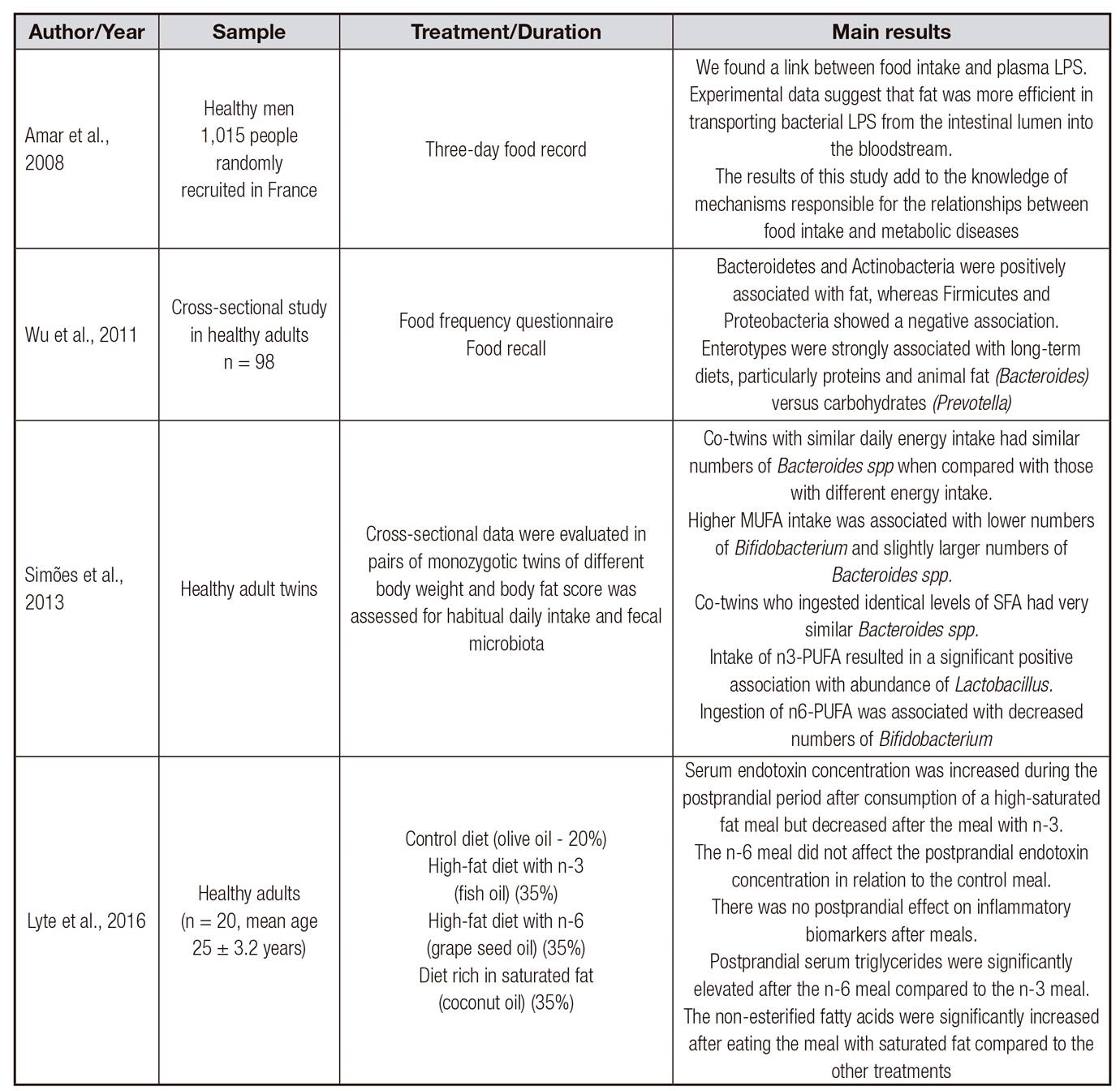
LPS: lipopolysaccharide; MUFA: monounsaturated fatty acids; PUFA: polyunsaturated fatty acids; SFA: saturated fatty acids.
Theoretically, dietary fat that reaches the colon can be metabolized by bacteria; cholesterol is degraded to form the metabolite coprostanol that is excreted in the feces 37. In addition, high-fat diets can stimulate the production of bile acids, which have antimicrobial activity and eventually select the species capable of metabolizing bile acids in the intestine 1. However, the use of fat as a source of energy for the growth of bacteria remains unclear, since fat metabolization is not performed in anaerobiosis, a condition common to most of the bacteria that inhabit the human intestine 38.
The influence of the diet rich in fat was analyzed in a study with mice fed a hyperlipid diet for three months, which showed a decrease of Bacteroidetes and increase of Firmicutes, Proteobacteria and Actinobacteria 29. Although long-term observations concisely present changes in the microbiota, it has been shown that short-term (five weeks) interventions are also capable of modifying the microbial ecosystem of mice 25. In addition, it has been reported that the diversity and composition of the microbiota, after being altered by the high-fat diet, can be reestablished with the intake of low fat diet.
Increased phylum Firmicutes was observed in wild-type mice receiving a high fat diet, and reduction of Bacteroidetes in genetically obese (ob/ob) resistant leptin mice 30. Thus, the authors suggested that a hyperlipid diet rather than the obese ob/ob genotype exerted a greater influence on the composition of the microbiota, which was also observed in another study 29. Thus, they emphasize that the microbiota changes observed in mice fed a high fat diet were probably attributable to changes in diet.
Similarly, the provision of a high-fat diet caused a general decrease in microbiota diversity and an increase in the ratio Firmicutes:Bacteroidetes in several studies with mice 25,32,39. Based on these findings, it was suggested that the microbiota of the obese has metabolic pathways that are highly efficient in extracting energy and favoring lipogenesis. This fact was supported by a study that, when transplanting the microbiota from obese mice to germ free mice, presented higher fat gain than animals transplanted with microflora from lean mice 40.
Despite reports of high concentrations of short chain fatty acids in the stools of obese individuals, these changes were not associated with a higher proportion of Firmicutes 41. This result generated research on the characteristics of the obesogenic microbiota, since it seems unlikely that only Firmicutes:Bacteroidetes are the only ones involved in the pathogenesis of obesity 30.
In a study with pairs of monozygotic and dizygotic twins, concordant for thinness or obesity and their mothers, Turnbaugh et al. 42 demonstrated that the microbial ecosystem is shared among family members, but that each individual has a specific bacterial composition. The authors also reported that obesity is associated with changes in phylum level in the microbiota, reduction of bacterial diversity and alteration of genes and metabolic pathways. A greater abundance of Actinobacteria, added to the Firmicutes phylum is reported, suggesting that the action of other phyla in the mechanisms that involve the microbiota and obesity interaction should be investigated.
In addition, mice fed a high-fat diet showed an increase in Clostridia XIV and Enterobacteriales and a decrease in Bifidobacterium spp 43. Similar results were also observed in the analysis of the microbiota after ingestion of a hyperlipic diet rich in palm oil and lard 33. Both diets were associated with increased relative abundance of Clostridiales spp and decrease of Bacteroidal.
The reduction in gram-positive bacteria such as bifidobacteria directly and negatively affects the integrity of the intestinal membrane, since they are responsible for maintaining and improving the intestinal barrier function, preventing the passage of bacteria and toxins 15. The same function can be attributed to the bacteria Akkermansia muciniphila, belonging to the phylum Verrucomicrobia, associated with stimulation of the immune system with anti-inflammatory properties 44. The increase in the abundance of Clostridiales spp may be related to mechanisms of metabolic pathways of cholesterol and levels of bile acids 33.
Some studies have reported that the fatty acid profile is also able to modulate the composition of the intestinal microbiota and contribute to the induction of low intensity inflammation (Table 3) 23,35,39,43. However, this subject requires more research and more in-depth knowledge 45.
Table III. Review of studies investigating the dysbiosis induced by different types of fats in animals

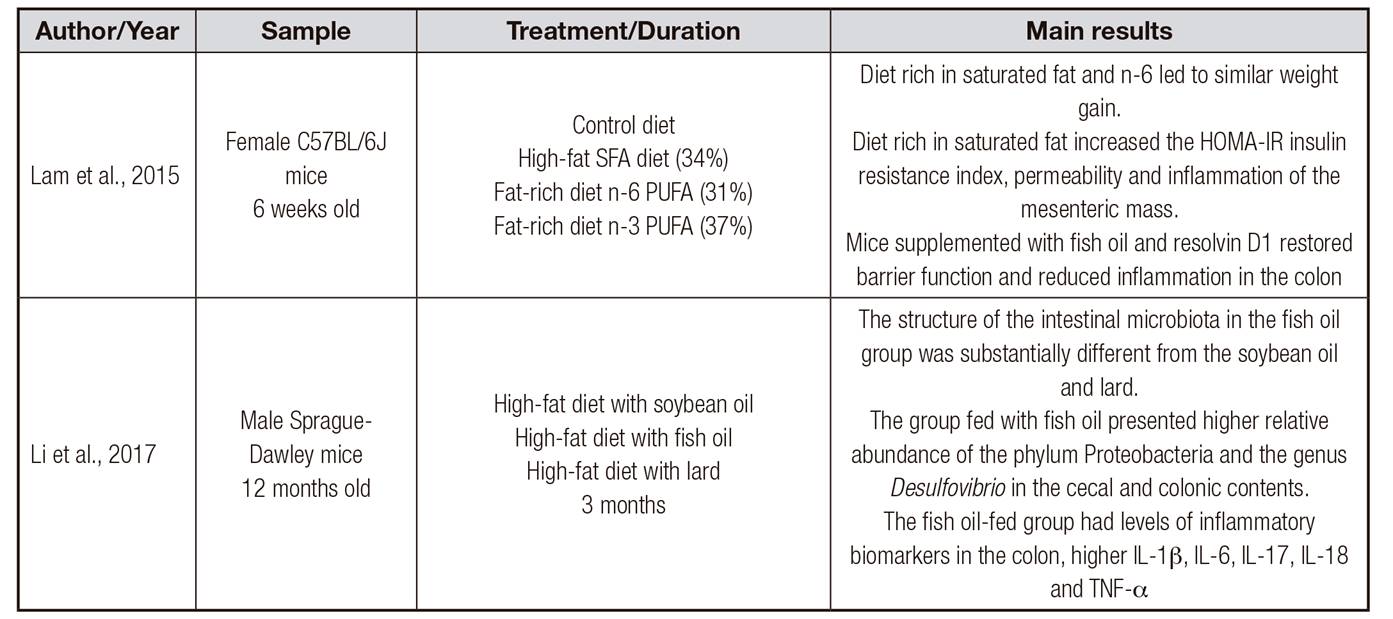
CHO: carbohydrate; LIP: lipids; PTN: protein; MUFA: monounsaturated fatty acids; PUFA: polyunsaturated fatty acids; SFA: saturated fatty acids; LPS: lipopolysaccharide; EPA: eicosapentaenoic acid; DHA: docosahexaenoic acid; HOMA-IR: insulin resistance index; IL-1β: interleukin-1β; IL-6: interleukin-6; IL-17: interleukin-17; IL-18: interleukin-18; TNF- α: tumor necrosis factor alpha.
The impact of different types of fats on the host's health, metabolism and microbiota was analyzed 23. Using mice fed a high-fat diet containing palm oil, olive oil, safflower oil and flaxseed oil/fish for 16 weeks compared to mice fed a low fat diet, it was found that mice populations fed palm had relatively lower populations of Bacteroidetes at the phylum level, compared to olive oil diet; this in turn showed an increase in the family Bacteroidaceae. However, mice fed flax/fish oil showed a significant increase in the concentrations of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) and in the intestinal abundance of Bifidobacterium. The findings suggest that the impact on the microbiota is due to both changes in the quantity and quality of fat ingested and that fats such as flaxseed oil positively impact the microbial ecosystem of the host.
Similarly, changes in the intestinal microbiota of humans were different according to the intake of different fatty acids, monounsaturated fatty acids (MUFA), omega-3 polyunsaturated fatty acids (PUFA) and omega-6 PUFA. While consumption of MUFA and omega-6 PUFA was inversely associated with an increase in the number of Bifidobacterium, an increase in omega-3 PUFA intake was directly associated with a higher number of bacteria in the Lactobacillus group 35.
In addition, in a study with middle aged (12 months) mice fed with saturated fat (lard), monounsaturated (soybean oil) and polyunsaturated (fish oil), different responses in the microbiota were observed. The group fed with fish oil showed higher relative abundance of the Proteobacteria phylum and the genus Desulfovibrio in the cecal and colonic contents. On the other hand, the diet based on saturated fatty acids conferred a higher Firmicutes:Bacteroidetes and more abundant presence of Verrucomicrobia and Tenericutes 46.
HIGH-FAT DIETS IN LIPIDS AND METABOLIC ENDOTOXEMIA
Obesity, diabetes and insulin resistance are associated with a low intensity systemic inflammation caused by multiple factors and whose triggering agents are not fully elucidated. However, the intestinal microbiota has been largely associated with this scenario 8,24. This evidence was obtained from studies that demonstrated the existence of endotoxemia, the passage of bacterial lipopolysaccharide (LPS) into the bloodstream during consumption of fat-rich diets by mice 47,48. This effect has also been confirmed in human studies 17,49 50 51-52,53,54,51.
Naturally the microbiota is a reservoir of LPS, since it is one of the components of the outer cell wall of gram-negative bacteria, and it is possible to detect more than 1 g of LPS in the intestinal lumen. Therefore, under normal conditions LPS is not harmful to the host and appears to be involved in immune regulation, such as increased phagocytic capacity, lymphocyte proliferation and lymphokine secretion. However, in situations of dysbiosis, which can be caused by the type of diet consumed, the transfer of LPS to the circulatory system can be increased and thus generate what we call metabolic endotoxemia 15.
Diet plays an important role in the regulation of endotoxemia. This evidence is supported by the fact that an increase in plasma LPS occurs in mice fed high-energy diets, whether rich in carbohydrates or fats, for four weeks. However, the high-fat diet proved to be more efficient in favor of LPS transfer from the intestinal lumen to the bloodstream 48.
An increase in postprandial serum endotoxin concentration was observed in healthy adults on high-fat diets, especially those on a saturated fat diet, when compared to subjects who received diets high in polyunsaturated fats 51. Similarly, men who received high-lipid meals also had elevated postprandial LPS levels when compared to fasted individuals 50. A possible explanation for this fact is the ability of LPS to be incorporated into micelles, absorbed and added to chylomicrons, due to the presence of a fraction insoluble in its molecular structure 52.
In addition, there may be an increase in local pressure and loosening of junctional complexes between enterocytes, or even basement membrane rupture, due to excess chylomicrons generated from the hyperlipidic diet. After the lesion caused during fat absorption the intestinal barrier may be compromised, increasing intestinal permeability, mainly to LPS 15.
Thus, consumption of high-fat diets leads to increased intestinal permeability and reduced expression of genes encoding tight or tight junction proteins, such as claudina-1, caludina-3, occludin, and junctional adhesion molecule 1 53. This is due to the regulation of permeability by mast cells, through the secretion of mediators such as tumor necrosis factor α (TNF-α), IL-1b, IL-4 and IL-13, receptor-2, favoring LPS translocation 54.
The fatty acid profile of the ingested lipid diet has been shown to be an important modulator of the microbiota, triggering endotoxemia 15. In a study with mice fed omega-6-rich meals, they exhibited conditions that characterized elevated levels of metabolic endotoxemia and low-intensity inflammation 55, while mice fed omega-3 fatty acids had decreased LPS production and permeability, with a significant reduction in metabolic endotoxemia.
In another study, it was observed that diet rich in fats (45% of lipids), when compared to control diet (13% of lipids), increased adiposity and plasma levels of lipopolysaccharide binding protein (LBP) ingestion for three and six weeks. However, with only one week of ingestion, an immediate but reversible increase in paracellular permeability and decreased expression of IL-10 was observed. In addition, a reduction of the abundance of genera within the class Clostridia in the ileum was observed 32.
Increased dietary fat content may influence phylum Actinobacteria, which may reduce the number of gram-positive Bifidobacterium species, considered to be beneficial. This may lead to an increase in LPS plasma concentrations, induction of low grade inflammation and maintenance of obesity 56. Likewise, inflammation can also occur with the increase of gram-negative bacteria, for example Desulfovibrio, capable of producing endotoxins, reduce sulfate to H2S and impair the intestinal barrier 20.
In addition, habitual intake of saturated fatty acids, derived from diets rich in fats and calories, can directly stimulate TLR4 cells. Alternatively, there is also a higher concentration of LPS by the increase of gram-negative bacteria, which can stimulate TLR4 and induce the expression of several cytokines, resulting in a state of low intensity inflammation and insulin resistance. In addition, increasing concentrations of circulating fatty acids may further increase nitric oxide production and decrease insulin sensitivity due to impaired lipoprotein lipase (LPL) activity and increased lipolysis 4.
An issue that deserves to be highlighted is the increase in bile production caused by the ingestion of high-fat diets, which acts selectively in relation to colonization of the intestine, being important in the modulation of the microbiota and in the role that it exerts on the permeability and production of endotoxins 57.
CONCLUSION
There is increasing scientific evidence that high-fat diets can modulate intestinal microbiota composition, enhancing LPS uptake and affecting mucosal integrity, resulting in metabolic endotoxemia. Installed endotoxemia is a causal factor of subclinical inflammation related to several chronic diseases and, as a result of this, it is essential to know the real impact of hyperlipidic diets on the microbiota.

