










 Custom services
Custom services
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El desarrollo de la nutrigénetica y la nutrigenómica ha establecido una relación entre los hábitos alimenticios y las enfermedades crónicas, ya que se ha demostrado que los nutrientes procedentes de la dieta interactúan con los genes 1. Los genes son las unidades funcionales y físicas de la herencia que pasa de padres a hijos 2. La expresión génica se refiere al proceso por el cual un gen se activa en una célula para elaborar RNA y proteínas y puede medirse al observar el RNA o la proteína elaborada con el RNA, o lo que la proteína hace en una célula 2. Los nutrientes, según la Organización Mundial de la Salud (OMS), son factores dietéticos de carácter orgánico e inorgánico contenidos en los alimentos y que tienen una función específica en el organismo. Los nutrientes principales en una dieta son, por un lado, los macronutrientes, como proteínas, hidratos de carbono simples y complejos, grasas y ácidos grasos, y por otro, los micronutrientes u oligonutrientes: vitaminas y minerales 3).
El contenido de proteínas en la dieta provoca concentraciones elevadas de aminoácidos (AA) y hormonas. Los AA son compuestos orgánicos que se combinan para formar proteínas 4 y las hormonas son los mensajeros químicos del cuerpo, que viajan a través del torrente sanguíneo hacia los tejidos y órganos 5. Todo ello provoca efectos que alteran la regulación de ciertos genes, tales como SNAT2 6 y AgRP 7. El hígado y el riñón participan en la regulación de los niveles de AA plasmáticos y en la síntesis de glucosa a partir del exceso de AA 8,9.
La literatura científica acumula evidencias sobre la influencia de AA en la expresión génica. Sin embargo, existe discrepancia en la información generada por la comunidad científica, ya que los modelos de estudios son variados y realizados en diferentes especies de animales. Por ello, el objetivo de este artículo es conocer la influencia de los aminoácidos provenientes de la dieta en la expresión de genes, mediante una revisión bibliográfica.
MATERIAL Y MÉTODOS
La revisión sistemática se realizó según los criterios establecidos por la declaración PRISMA 10), considerando, además, la reducción razonada de Maeda 11. Se efectuó una búsqueda en bases de datos de Scielo, PubMed, Medline, US National Library of Medicine National Institutes of Health (NCBI), Springer, Scopus, Science Direct y Elsevier de artículos publicados en inglés y español. El periodo de revisión tuvo como límite el 25 de mayo de 2018. Los términos de búsqueda fueron: "expresión génica", "aminoácidos", "síntesis de proteínas", "glutamina", "arginina" y "mTOR". La procedencia de las publicaciones fue de diversos países como España, Francia, Suecia, Chile, Alemania, Australia, Corea, China, Inglaterra y Estados Unidos.
CRITERIOS DE INCLUSIÓN
Se tomaron en consideración criterios de selección fundamentales (título, resumen y resultados) para clasificar los artículos como sigue:
Artículos de relevancia para el desarrollo del tema, que incluyeran los términos de búsqueda; estudios genéticos en animales.
Factor de impacto (0.500-5.000), obtenidos de los principales índices de citación (Journal Citation Reports, JCR; Scimago Journal & Country Rank, SJR).
Artículos publicados en revistas indizadas.
Estudios preferentemente enfocados a la nutrición.
RESUMEN DE LA BÚSQUEDA
Tras la primera exploración se encontraron 105 referencias relacionadas con los términos de búsqueda, de las cuales solo se seleccionaron los estudios que cumplían con los criterios de inclusión. Se analizaron 71 resúmenes, de los que se dio lectura a 30 textos completos que fueron seleccionados para la revisión
(Fig. 1). Además, se consultaron seis libros relacionados con la nutrición, los aminoácidos y la expresión génica. También se incluyeron cuatro referencias para definir los conceptos que forman parte de la introducción y dos sobre la metodología de revisión (PRISMA y Maeda). La tabla 1 muestra un resumen de los principales artículos seleccionados para la revisión.


INFLUENCIA DE AA EN LA EXPRESIÓN GÉNICA
Algunos genes se regulan en varios pasos que incluyen transcripción, procesado postranscripcional, exportación nuclear y traducción de los mRNA maduros. Los AA pueden tener una influencia indirecta en la modulación al servir como señales en la transcripción debido a que se ha demostrado que las células pueden detectar variaciones en los niveles de AA y responder por mecanismos como el control de transcripción, la estabilización del RNA mensajero y el incremento o la disminución del inicio de la traducción 12.
Los mecanismos epigenéticos más importantes de expresión génica incluyen la modificación de proteínas histonas, que definen las regiones del cromosoma en las que el ADN se libera temporalmente para permitir el acceso de factores de transcripción 12. Las histonas se modifican mediante metilación, acetilación y fosforilación. La región promotora es amplia en eucariotas. En ella se encuentran repeticiones de secuencias de nucleótidos de citosina unida a guanina, CpG (dentro de la misma cadena, marca puntos en los que una metiltransferasa puede agregar un grupo metil a la citosina en el carbono número 5). Cuando existe una mayor metilación, los factores de transcripción no reconocerán la región promotora. Estos procesos son de vital importancia para la regulación de la expresión génica y las funciones fisiológicas 13. En la acetilación de las histonas, el ADN es disociado de sus histonas y la transcripción procede. Por el contrario, la metilación del ADN y la desacetilación de histonas provocan que el empaquetamiento del ADN aumente y se silencie la expresión genética 14.
La transcripción de un gen a un mRNA se efectúa mediante la RNA polimerasa. Este proceso puede ser regulado por los AA a través de la activación de factores de transcripción, de C/EBP (CCAATl, enhancer binding protein) y de la regulación de secuencias específicas en el promotor 15.
Existen varios genes que codifican para proteínas ribosómicas que están regulados por la disponibilidad de AA, como los genes S25 y L17, que codifican proteínas para la subunidad ribosómica 60S. Entre los represores más efectivos para la inducción del L17 y la asparagina sintetasa (AS) se encuentran la glutamina (Gln), la asparagina (Asp) y el aminoisobutírico 16.
La Gln y Arg son dos AA sumamente investigados en la regulación de la expresión de genes.
GLUTAMINA
La Gln es considerada como la principal fuente de energía de células epiteliales (enterocitos del yeyuno y de linfocitos) y de células del sistema inmune. Este AA juega un papel importante en el transporte de nitrógeno, síntesis proteica, actuando como un sustrato de la amoniogénesis renal y neoglucogénesis hepática 17. La suplementación de Gln ha sido efectiva para reducir la morbimortalidad de los pacientes catabólicos y proteger contra fenómenos de lesión del estrés oxidativo 18.
La Gln en la dieta incrementa la expresión de los genes (120-124%) que son necesarios para el crecimiento celular y de remoción de oxidantes. Reduce la expresión (34-75%) de genes que promueven el estrés oxidativo y la activación inmune 19,20. La adición de Gln a fórmulas de uso enteral y parental aumenta sus concentraciones en sangre, mejora el balance nitrogenado y disminuye la incidencia de infecciones y la duración de estancias hospitalarias. Asimismo, reduce la muerte celular provocada por shock térmico, induciendo específicamente la hsp 70 y 72 en células intestinales, mientras que la privación induce apoptosis de enterocitos. Sin embargo, se desconocen los mecanismos moleculares íntimos de modulación en la expresión de estos y otros genes 16.
Por otra parte, se ha observado experimentalmente la implicación de la Gln en la expresión de C/EBP, responsable de la unión de proteínas y SLC1A5, necesario para el transporte de AA y Gln, en células de hepatoma 17. Además, la Gln puede contribuir a diversas acciones relativas a la proliferación y supervivencia celular, tales como la síntesis de proteínas y nucleótidos, el descenso de la proteólisis y de la acción moduladora sobre ciertos factores de crecimiento como el IGF, la GH y la inhibición de la apoptosis 14. Los efectos inmunosupresores de la Gln han sido estudiados y se ha demostrado que modula la acción de varios factores de transcripción, especialmente el NF-kB 21.
ARGININA
Fisiológica y nutricionalmente, la L-arginina (Arg) es de vital importancia, ya que estimula la secreción de hormonas que están involucradas en el crecimiento y en el metabolismo, tales como la insulina y el glucagón 21. Adriao y cols. 22 investigaron el papel de la Arg en pituitarias de rata y células GH3, observándose un aumento en expresión génica de GH una hora después del tratamiento con este AA. En otro estudio similar, la L-Arg indujo significativamente la secreción de la hormona GH e IGF-1 de las células GH3 y HepG2, respectivamente 23.
La L-arginina es de vital importancia. En un estudio se analizó la "viabilidad" de células β, es decir, la pérdida de función celular, y se encontró una disminución de la misma de un 100% hasta un 61% en ausencia de L-Arg. Si, además, las células se exponían a citoquinas proinflamatorias, el valor disminuía del 100% al 41%. Por otra parte, la L-Arg también es precursora natural del óxido nítrico (NO), que funciona como vasodilatador en el endotelio vascular 24).
Una adecuada suplementación con Arg reduce los niveles de mRNA para proteínas de unión de ácidos grasos y proteína fosfatasa 1B. Incrementa la expresión de la glutatión sintetasa de insulina (GSS) como factor de crecimiento II, proteína quinasa AMPK (activated protein kinase). En estudios bioquímicos, Jobjen y cols. 25 demostraron una disminución en el estrés oxidativo de células de ratas obesas WAT (white adipose tissue), cuando se les administraba Arg. La Arg activa la vía de señalización mTOR (proteína quinasa diana de rapamicina de mamíferos) en enterocitos, que estimula la síntesis proteica, la migración celular y la reparación intestinal, reduciendo los procesos inflamatorios 21.
CONTROL DEL INICIO DE LA TRADUCCIÓN POR AA
La traducción genética es regulada por un conjunto de mecanismos que actúan en la fase de iniciación, elongación y terminación 26. Los AA pueden regular la síntesis de proteínas a través de cambios en eIF2B, fosforilación de 4E-BP1 y de proteínas S6 27.
MODULACIÓN DE LA ACTIVIDAD DEL EIF2B
El inicio de la traducción se desarrolla en diferentes etapas (Fig. 2):
Activación del complejo eIF2 por medio de una molécula de GTP para formar el eIF2-GTP.
El eIF2-GTP se unirá al complejo met-tRNA y a la subunidad 40 S del ribosoma para la formación del complejo 43 S (Met-tRNA-eIF2-GTP-40 S) y para seguir con el proceso de síntesis de proteínas.
El GTP del complejo 43 S es hidrolizado para formar GDP. Después de cada ciclo de iniciación, el eIF2 se libera como un complejo binario unido a GDP y eIF2-GDP regresa a su estado de inactivación.
Con el objetivo de volver activar el eIF2, el GDP se intercambia por GTP y se forma de nuevo eIF2-GTP, que funcionará en otro ciclo de iniciación. Este intercambio de nucleótidos de la guanina está catalizado por otro factor de iniciación, el eIF-2B, que regula el primer paso de iniciación 27,28.
Existen dos mecanismos que pueden regular la actividad del eIF-2B: la fosforilación de la subunidad α del eIF2 y la fosforilación de la subunidad ε del eIF-2B (Fig. 2). El eIF2α es uno de los dos puntos de control para la iniciación de la síntesis de proteínas en las células eucariotas 26. Cuando existe una disminución en el consumo de AA, el eIF-2α se fosforila en la serina 51 gracias a una enzima llamada general control non-depressing kinase-2 (GCN2 kinasa). La fosforilación del eIF-2α por GCN2, provoca que se forme un complejo con el eIF-2B (eIF-2α-eIF-2B), lo que impide la unión del Met-tRNA-eIF2-GTP a la subunidad 40S del ribosoma. El resultado es la inhibición del inicio de la traducción, interrumpiéndose la síntesis de proteínas 16,26. También existe un mecanismo en donde la activación de la GCN2 implica la acumulación de tRNA desacilados. Los elementos de respuesta a aminoácidos (amino acid response elements [AARE]) pueden regular la fase de iniciación de la síntesis de proteínas a través del factor de iniciación eIF-2α 29.

FOSFORILACIÓN DEL COMPLEJO 4E-BP1
Existe un segundo paso en la iniciación de la traducción donde se une un mRNA al complejo de preiniciación 43 S. Este proceso se efectúa por un complejo de factores de iniciación denominados eIF-4F, los cuales están integrados por una helicasa de RNA (eIF-4A), una proteína que se une a la caperuza m7GTP del extremo 5' del mRNA (eIF-4E), la proteína que sirve de andamio (eIF-4G) para unirse con eIF-4A y la proteína de unión a la cola de poli-A (poli A binding protein [PABP]) 21. Estas tres proteínas (eIF-4A, eIF-4E y eIF-4G) se adhieren al complejo de iniciación 43 S y este, a su vez, a la cadena de mRNA 27.
El ensamblaje del complejo eIF-4F está regulado en parte por la asociación del eIF-4E con las denominadas proteínas de unión al eIF-4E (4E-BP1) 30) (Fig. 3). El sitio de unión de 4E-BP1 con eIF-4E se solapa con eIF-4G; de este modo, se pueden unir individualmente al eIF-4E, pero no ambos al mismo tiempo. Entonces, la unión del eIF-4E a la proteína 4B-BP1 impide la asociación del mRNA al ribosoma; esto ocurre únicamente cuando 4E-BP1 se encuentra hipofosforilada. De forma contraria, cuando se hiperfosforila la 4E-BP1, se estimula el ensamblaje del complejo eIF-4E con eIF-4G 16,31.
Si existe una privación de AA en la dieta (especialmente Leu),

Figura 3 Activación de mTOR mediante la privación de AA de la dieta, fosforilación del 4E-BP1 y de la proteína S6. Adaptado de Fafournoux y cols. (27).
la mTOR se activa y causa la fosforilación de 4E-BP1. La mTOR (target of rapamycin) es otro tipo de quinasa cuya función principal es coordinar la disponibilidad de nutrientes con el crecimiento celular. Asimismo, fosforila las proteínas que se requieren para la unión del mRNA con la subunidad 40 S del ribosoma y la proteína ribosómica S6 (rpS6, ribosomal protein) 21,32 (Fig. 2).
FOSFORILACIÓN DE LA PROTEÍNA S6
La S6 es una proteína ribosomal y su fosforilación se realiza directamente por una proteína llamada S6 kinasa I (S6KI), cuya actividad es a su vez regulada por la fosforilación catalizada por la mTOR, que para activarse requiere de una adecuada disponibilidad de AA y de la presencia de insulina 21,33. La fosforilación de S6 aumenta la traducción de un conjunto de mRNA específicos, los cuales codifican para proteínas ribosómicas, implicadas en la traducción. La mTOR controla la síntesis de rRNA y proteínas ribosómicas, es decir, la biogénesis de los ribosomas 21,34. La propia traducción de mRNA mTOR podría estar regulada por la fosforilación de la rpS6 16.
AMINOÁCIDOS EN LA TRANSCRIPCIÓN DE GENES
El control de AA en la transcripción de genes ha sido estudiado utilizando los mecanismos moleculares que involucran a los genes CHOP (C/EBP homologous protein) y el gen de la asparagina sintetasa (AS).
GEN CHOP
El gen CHOP es una proteína nuclear relacionada con la familia de los factores de transcripción C/EBP (CCAATl), que dimeriza con otros miembros de su familia y está involucrada en la apoptosis celular 27. Por otra parte, en el promotor del gen CHOP se ha identificado un elemento de respuesta a AA denominado amino acid response element (AARE), capaz de inducir la expresión en respuesta al ayuno total de AA 16. Existe una similitud entre la secuencia de AARE (5'-ATTG-CATCA-3') con los sitios cis específicos de las familias de factores de transcripción C/EBP y ATF/CREB. Entre estos factores, solo el ATF2 y el ATF4 están involucrados en la regulación dependiente de los AA por el AARE 1,29.
El ATF2, es un factor de transcripción cuya activación génica se regula vía fosforilación de dos restos de Thr y uno de Ser. La supresión de AA, especialmente de Leu, induce la fosforilación del ATF2 en células humanas, necesaria para que el gen CHOP se exprese.
El ATF4 es el primer regulador de la expresión del gen CHOP al inicio de la traducción. Al igual que el ATF2, este es activado o inducido por la falta de AA en la dieta, particularmente por Leu, y puede interaccionar in vitro con el AARE del gen CHOP provocando la activación génica 16,35).
GEN DE LA ASPARAGINA SINTETASA (AS)
Los AA, pueden tener influencia sobre la expresión del gen AS, lo que ocasiona un incremento de la proporción de transcripción y de estabilidad del RNA mensajero. Para la síntesis de la Asn y Glu a partir de Gln y Asp se requiere una enzima llamada asparagina sintetasa y ATP 36. La transcripción del gen AS aumenta en respuesta a una falta de AA o glucosa. Así, los niveles de tRNA-Asn disminuyen cuando decrece la concentración de Asp, mientras que la actividad y los niveles de mRNA AS aumentan. De este modo, la Gln y otros AA en menor medida intervienen en la represión del gen AS. En el análisis de tejidos de páncreas se ha observado una mayor expresión del gen AS y es recurrente en diferentes especies como humanos, pájaros y roedores 36.
EXPRESIÓN GÉNICA DE FACTORES DE CRECIMIENTO Y AA
Un adecuado crecimiento, especialmente en niños, requiere de una compleja interacción entre diversos factores genéticos, hormonales y nutricionales. Una parte de este control es realizado gracias al factor de crecimiento IGF-I e IGF-2, abundantes en la circulación y producidos por la mayoría de los tejidos del cuerpo 37. La actividad biológica de los IGF es modulada por proteínas de unión llamadas IGF binding proteins (IGF-BP) 37. La expresión del gen que codifica para el factor de crecimiento análogo a la insulina (insulin like grow factor [IGF-I]) en el ser humano está regulada por la disponibilidad de algunos AA. Es decir, los niveles de mRNA de IGF-I hepáticos y los niveles de IGF-I plasmáticos se correlacionan con la velocidad de crecimiento y disminuyen cuando existe un déficit de nutrientes. Por el contrario, las concentraciones de mRNA para las proteínas de fijación de IGF-I (IGF-BP-I) aumentan 27. La nutrición influye en la biosíntesis y la secreción hepática de IGF e IGFBP, provocando una afectación en órganos como el sistema inmune y el intestino. El IGF-I eleva la proliferación de células T y B 9.
Jousse y cols. 37 demostraron que la falta de AA induce la expresión génica de IGF-BP-I por la disminución de Arg, Cys y todos los AA esenciales, provocando afecciones significativas en los niveles de mRNA del IGF-BP-I en una línea de células de hepatoma (HepG2). Los datos obtenidos revelaron que la limitación de Leu induce fuertemente la expresión génica de IGFBP-I, sin afectar la expresión de IGF-I y IGF-II en células humanas HepG2.
Por otra parte, Passos de Jesús y cols. 38 encontraron que la suplementación con Pro y Gln en ratas desnutridas mejoró el contenido total de RNA en el tejido hepático remanente. La administración de AA aumentó la expresión del gen del factor de crecimiento de hepatocitos (hepatocyte growth factor [HGF]) después de la hepatectomía parcial en roedores desnutridos, teniendo mayor efecto Pro que Gln.
OTROS EJEMPLOS DE GENES REGULADOS POR AA
En la tabla 2 se muestra cómo aumenta o disminuye la expresión génica de algunos genes.
En una investigación sobre AA que actúan sobre las neuronas hipotalámicas para regular el comportamiento de la alimentación y la homeostasis energética, se utilizó una dieta baja en proteínas (10% de las calorías) en ratas. Los resultados exhibieron un aumento en la ingesta de alimentos y en la expresión del gen hipotalámico agouti-related protein (AgRP). Después, mediante una inyección intracerebroventricular directa, una mezcla de aminoácidos (RPMI 1640) o Leu (1 g) fue suministrada, estimulando una disminución en la ingesta de alimentos de los roedores durante 24 h. Estas observaciones indican que los AA pueden actuar dentro del cerebro para inhibir o aumentar la ingesta de alimentos y que la expresión del gen AgRP puede contribuir a este efecto. Sin embargo, los mecanismos de señalización que median estos efectos no están del todo claros 7.
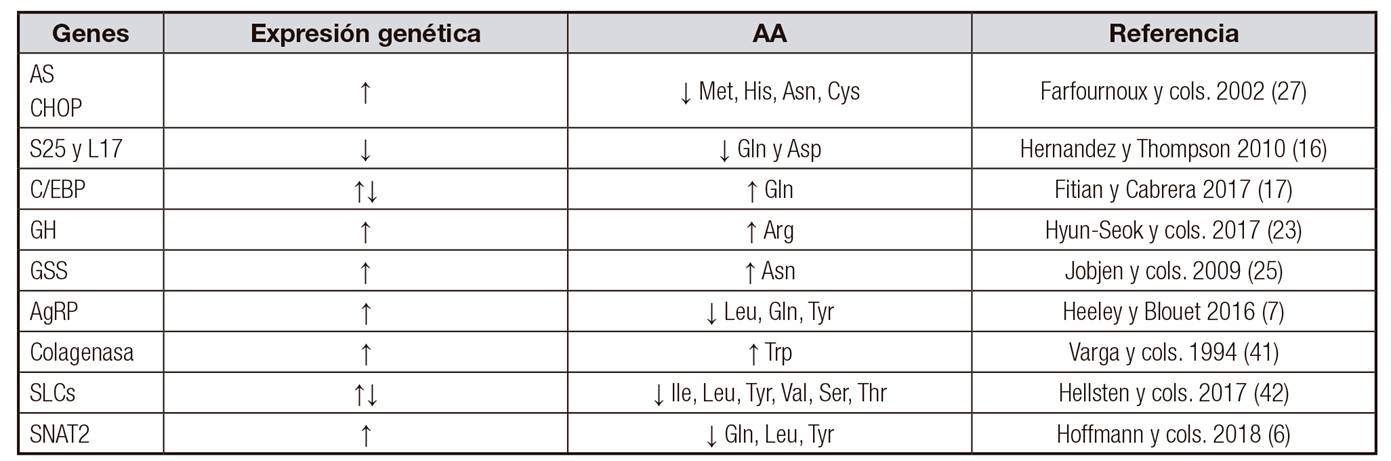
La capacidad de distintos AA para estimular la secreción hormonal depende de cada hormona y especie. Los AA (Leu, Gly, Ser, Ala) aumentan la secreción de insulina cuando se inyectan en animales de experimentación 9. Esto se debe, probablemente, a que la insulina favorece la entrada de AA a la célula y su incorporación a las proteínas, además de estimular la síntesis e inhibir el catabolismo de proteínas.
La secreción de hormona de crecimiento (GH) es controlada por su factor hipotalámico (GHRF) y somatostatina. Estudios experimentales han demostrado que el ayuno reduce la concentración del mRNA del GHRF, pero no afecta al mRNA de la somatostatina 39.
En mujeres embarazadas, una dieta rica en proteínas ocasiona la expresión de enzimas como la ribonucleasa (Rnasa), tiorredoxina reductasa (TR) y lactato deshidrogenasa (LDH), entre otras. De forma contraria, con una dieta materna baja en proteínas se expresan mayoritariamente las enzimas glutatión S transferasa (GST), ornitina carbamoiltransferasa (OCT) y aspartato transaminasa (AST) 1.
En células humanas, el AA L-triptófano (Trp) fue un potente inductor de la expresión de la colagenasa (enzima encargada de degradar glucógeno, que además contiene zinc) a nivel de transcripción, donde el incremento de mRNA de colagenasa fue reversible, dependiente del tiempo y de la concentración del L-triptófano 40,41.
Los SLCs son una gran familia de transportadores de proteínas en mamíferos y su expresión génica se ve afectada notablemente si existe una privación de AA como Ile, Leu, Tyr, Val, Ser y Thr, lo cual implica un efecto en la capacidad de los SLCs para regular las concentraciones de AA intracelulares y, además, detectar alteraciones en los niveles de AA extracelulares. Un estudio reciente determinó que la expresión génica de los SLCs se incrementaba en células hipotalámicas N25/2 de ratones después de la privación de AA como Ile, Leu, Tyr, Val, Ser, Thr y Trp 42.
El SNAT2 es otro transportador de AA que interviene en la absorción celular de AA neutros acoplados a Na+ y está regulado mediante la activación de mTORC1 (mammalian target of rapamycin complex 1). La disminución de concentraciones de AA extracelulares (Gln, Leu, Tyr) induce un incremento en la expresión del SNAT2 como respuesta a una concentración disminuida de AA; en tal caso, también se debe considerar una afectación en la activación y regulación de mTORC1, lo cual a su vez afectaría el proceso de síntesis de proteínas y crecimiento celular 6.
CONCLUSIÓN
La síntesis de proteínas en células de mamíferos es controlada por dos vías de señalización, mTOR y vía sensible a aminoácidos (amino acid response [AAR]). La vía de mTORC se activa cuando la célula tiene niveles idóneos de AA y funciona como un sensor, a fin de mantener la síntesis de proteínas y el crecimiento celular. De forma contraria, la AAR se activa cuando la célula es privada o tiene bajos niveles de AA, lo que deriva en la inhibición de la síntesis general de proteínas. El control de los procesos concernientes a la transcripción, la traducción y las modificaciones postransduccionales modulados por los AA no ha sido muy estudiado, aunque en los últimos años se ha experimentado con AA tales como Gln, Asp y Leu en los mecanismos de transcripción génica. En este sentido, se ha observado el efecto de AA sobre la modulación de la cromatina provocando la metilación del DNA y la acetilación de histonas. Sin embargo, las acciones de AA en la modulación de la actividad del eIF2B, la fosforilación del complejo 4E-BP1 y la fosforilación de la proteína S6 son más evidentes. Por otra parte, los AA pueden regular la expresión de genes que codifican para factores de crecimiento (como el IGF-I en el ser humano) y hormonas (la secreción de insulina aumenta cuando se suministran Leu, Gly, Ser y Ala en animales de experimentación).
En resumen, los avances en nutrigenómica y nutrigenética proporcionan la oportunidad de relacionar nutrientes de alimentos con la salud. De esta manera, podrían realizarse intervenciones dietéticas oportunas seleccionando los nutrientes específicos para cada individuo en base a su carga genética, previniendo enfermedades en un futuro. Los AA, que actúan sobre las neuronas hipotalámicas para regular el comportamiento de la alimentación y la homeostasis energética, son un ejemplo de cómo se puede manipular la expresión de un gen para inducir o no la ingesta de alimentos, lo que ofrece una posibilidad de mejora en el tratamiento de pacientes con obesidad, diabetes o enfermedades del corazón.

