










 Custom services
Custom services
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
La acrilamida es un compuesto tóxico que se forma durante el sometimiento de alimentos ricos en carbohidratos a temperaturas elevadas, mayores a 120 °C. Los productos alimenticios derivados de ingredientes vegetales como papas y cereales tienden a contener las mayores cantidades de acrilamida; esto se debe principalmente a la presencia natural de compuestos como glucosa, fructosa y asparagina, involucrados en la formación de acrilamida (1). Los efectos neurotóxicos de la acrilamida en humanos han sido bien documentados, sugiriendo la capacidad para causar mutaciones genéticas. Asimismo, la recombinación inducida por el ADN puede interferir en el proceso de replicación genética, lo que favorece la formación de tumores (2). Por otra parte, las bacterias acidolácticas constituyen un grupo de bacterias grampositivas que forman parte importante de la microbiota intestinal debido a que desempeñan un papel trascendente en el buen funcionamiento del organismo y el mantenimiento de la salud (3,4). Además se sabe que las bacterias acidolácticas generan compuestos con actividad antioxidante que neutralizan moléculas tóxicas a nivel intestinal y sistémico (5), además de haber demostrado tener actividades antigenotóxicas y antimutagénicas en modelos in vitro e in vivo (6,7). El uso de las células no viables de bacterias acidolácticas, o sus componentes y/o extractos, podría tener ventajas de seguridad sobre la utilización de las bacterias completas ya que en los consumidores con sistemas inmunes desequilibrados o comprometidos existe el riesgo de traslocación, infección o aumento de la respuesta inflamatoria, mostrado por algunas cepas de BAL con potencial probiótico. Es por esto que el presente trabajo pretende evaluar el efecto de cepas de Lactobacillus y sus extractos intra y extracelulares contra la genotoxicidad y el estrés oxidativo causados por la acrilamida.
METODOLOGÍA
SOLUCIÓN DE TRABAJO DE ACRILAMIDA
Se preparó una solución de acrilamida disolviendo esta en solución de buffer de fosfatos salino (0,9 % NaCl en PBS 10 mM, pH = 7,4) para alcanzar una concentración final de 100 mM.
CULTIVO BACTERIANO
Se empleó una cepa de Lactobacillus casei Shirota aislada de un producto lácteo fermentado comercial, y una cepa de Lactobacillus reuteri NRRL B-14171 que pertenece a una colección de cultivos. Para cada cepa se realizaron dos subcultivos en caldo Man Rogosa y Sharpe (MRS) para obtener concentraciones celulares de 109 UFC/mL previo a cada experimento.
PREPARACIÓN DEL CONTENIDO INTRACELULAR DE L. CASEI SHIROTA Y L. REUTERI ATCC 14171
En breve, se tomó una alícuota (10 ml) de bacterias suspendidas en PBS a la que se le añadió 1 mg/ml de lisozima y se incubó a 37 °C durante 2 h; posteriormente, las células tratadas fueron sometidas a ultrasonicación (70 W) y recuperadas por centrifugación (3600 g, 10 min, 4 °C); el sobrenadante se mantuvo en refrigeración y oscuridad hasta su uso y los restos de bacteria se emplearon para obtener los extractos extracelulares.
EXTRACCIÓN DE PARED CELULAR Y ÁCIDOS TEICOICOS DE L. CASEI SHIROTA Y L. REUTERI NRRL B-14171
Los restos de bacteria obtenidos después de la ruptura para obtener el líquido intracelular fueron tratados con proteinasa K y lisozima para romper la estructura bacteriana. Finalmente, los fragmentos se trataron con dodecilsulfato sódico (SDS) y ácido etilendiaminotetraacético (EDTA). Los fragmentos que se obtuvieron se mantuvieron en congelación hasta su uso para obtener ácidos teicoicos y para realizar los ensayos de micronúcleos y del grado de oxidación. Por otra parte, se tomaron 10 g de los fragmentos de la pared celular aislada de las cepas a evaluar y se incubaron durante 24 h a 37 °C en 5 mL de ácido tricloroacético (TCA) al 10 % sin agitación. Posteriormente, los ácidos teicoicos se recuperaron por precipitación con 5 mL de acetona fría durante 24 h a 4 °C y se recuperaron los ácidos teicoicos por centrifugación (3200 x g, 10 min, 4 °C).
EVALUACIÓN DEL EFECTO ANTIOXIDANTE
La capacidad antioxidante se evaluó en los extractos extra e intracelulares de ambas cepas evaluadas mediante la técnica ABTS. En breve, se preparó una solución del radical ABTS (7 mM), la cual se dejó reaccionando con una solución de persulfato de potasio (2,45 mM) durante 16 h a temperatura ambiente y en obscuridad. Trascurrido el tiempo se almacenó a -80 °C en alícuotas de 1 mL hasta el momento de ser requerida. Una vez preparado el radical, se tomó una alícuota de la solución “stock” de ABTS y se ajustó la absorbancia a 0,07 ± 0,05 usando metanol absoluto, y se leyó a 750 nm en un espectrofotómetro UV-VIS (Multiskan Sky); adicionalmente, se preparó una curva de calibración a concentraciones de 5-300 µg/mL utilizando ácido ascórbico (1 mg/mL) como estándar para conocer las concentraciones de las muestras.
EVALUACIÓN DE LA CAPACIDAD ANTIOXIDANTE EN SACCHAROMYCES CEREVISIAE
La prueba de protección de la oxidación generada por H2O2 se evaluó empleando una cepa de Saccharomyces cerevisiae como modelo celular siguiendo el siguiente diseño experimental: 1) levadura + H2O2; 2) levadura; 3) bacteria completa + levadura + H2O2; 4) pared celular + levadura + H2O2; 5) líquido intracelular + levadura + H2O2; 6) ácidos teicoicos + levadura + H2O2. La metodología seguida fue la descrita por Oliveira y cols., 2021, en donde 0,1 g de Saccharomyces cerevisiae RC 212 (Lalvin bourgovinä) se colocaron en 5 mL de caldo YPD (Yeast extract, Peptone, Dextrose, Bioxonä) y se incubaron por 16 h a 28 °C con agitación a 180 rpm en agitador orbital. Transcurrido el tiempo se tomaron 100 µL, se pasaron a otro tubo con 5 mL de caldo YPD y se incubó el conjunto 16 h a 28 °C con agitación. De este último cultivo se colocaron 100 µL en tubos con 5 mL de caldo YPD y 100 µL de los extractos intra y extracelulares de ambas cepas de acuerdo con el diseño experimental descrito anteriormente, y se llevaron a incubación por 1 h; después se agregaron 7,5 µL de H2O2 y se incubaron por 1 hora más. De la mezcla anterior se prepararon diluciones seriadas en caldo YPD hasta 1 x 10-5 UFC y se sembraron por vaciado en placa de ágar YPD. Las placas se incubaron a 28 °C por 48 h; transcurrido el tiempo, se realizó el recuento en placa y los resultados se reportaron como UFC/mL de levaduras.
EVALUACIÓN DEL EFECTO ANTIGENOTÓXICO DE L. CASEI SHIROTA Y L. REUTERI NRRL B-14171
La prueba de formación de micronúcleos en los linfocitos se realizó de acuerdo con lo propuesto por Zamani y cols. (8) siguiendo el siguiente diseño experimental: 1) control (acrilamida); 2) acrilamida + bacteria completa; 3) acrilamida + líquido intracelular; 4) acrilamida + ácidos teicoicos; 5) acrilamida + fragmentos de pared celular. En breve, 0,5 mL de sangre humana se mezclaron con 4,5 mL de medio Roswell Park Memorial Institute (RPMI) 1640 suplementado con suero fetal de ternera al 20 %, penicilina 100 U/mL y estreptomicina 100 mg/mL, a lo cual se añadió fitohemaglutinina-M (1 mg/mL) para estimular el cultivo, incubándose la mezcla a 37 °C durante 72 h. A las 24 h del primer día, las células se trataron de acuerdo con el diseño experimental y, posteriormente, después de 20 h de exposición, se añadió citocalasina B (5 mg/mL) con el objetivo de bloquear la citocinesis celular; 28 h después se obtuvieron los linfocitos por centrifugación. Los linfocitos se sometieron a un tratamiento hipotónico suave con KCl (0,075 M) durante 5 minutos para luego fijarlos en un portaobjetos con una solución de metanol:ácido acético (3:1), repitiendo la fijación 2 veces. Finalmente, una vez fijada, la suspensión celular se tiñó con colorante de Giemsa al 10 % (pH = 6,8) durante 10 min. El porcentaje de micronúcleos se reportó por cada 1000 células observadas utilizando un microscopio de luz óptica.
MEDICIÓN DE LA PEROXIDACIÓN LIPÍDICA EN LINFOCITOS HUMANOS
La medición de la peroxidación lipídica se realizó en los linfocitos cultivados de acuerdo con el diseño experimental descrito en la sección anterior. El contenido de malondialdehído (MDA) se determinó por las sustancias reactivas del ácido tiobarbitúrico (TBA), de acuerdo con lo descrito por Zamani y cols. (8), con algunas modificaciones. En breve, se añadieron 0,25 mL de ácido sulfúrico (0,05 M) a los linfocitos obtenidos de cada grupo para, posteriormente, adicionar 0,3 mL de TBA (0,2 %) y colocarlos en un baño de agua hirviendo durante 30 min. Al finalizar el tiempo, se llevaron los tubos a un baño de hielo, se añadieron 0,4 mL de n-butanol a cada uno y se centrifugaron a 3500 g durante 10 min. Finalmente, se midió la cantidad de 1,1,3,3,-tetraetoxipropano (TEP) formado en cada muestra, midiendo la absorbancia del sobrenadante a 532 nm con un lector de ELISA y expresándolo como mmol/L de TEP.
ANÁLISIS ESTADÍSTICO
Los ensayos se realizaron por triplicado y los resultados se presentan como media ± desviación estándar. La significancia estadística (p < 0,05) se determinó mediante la prueba estadística ANOVA de una cola con una prueba de Tukey, mediante la cual se buscaron diferencias significativas en la capacidad antioxidante y el número de micronúcleos formados por cada 1000 células observadas al microscopio de luz óptica entre los diferentes tratamientos evaluados. Para el análisis estadístico se empleó el software estadístico Minitab 16.
CONSIDERACIONES ÉTICAS
Este estudio se apegará a lo señalado por la Declaración de Helsinki y lo dispuesto en la Ley General de Salud en materia de investigación. En todo momento se protegió la confidencialidad de la información y se recabó el consentimiento informado de los participantes en la investigación.
RESULTADOS
EVALUACIÓN DEL EFECTO ANTIOXIDANTE DE L. CASEI SHIROTA Y L. REUTERI NRRL B-14171
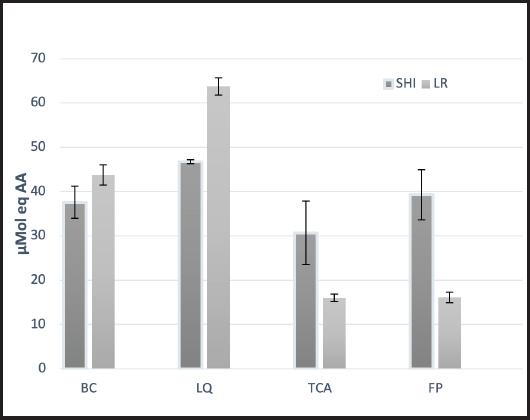
En la figura 1 se muestran los resultados referentes a la capacidad antioxidante de los extractos intracelulares y extracelulares de L. casei Shirota y L. reuteri NRRL B-14171 obtenidos mediante la técnica ABTS. De ambas cepas se obtuvieron valores que oscilaron entre 15,99 y 63,77 µME AA (micromoles de equivalente de ácido ascórbico). Al comparar la capacidad antioxidante de los extractos de ambas cepas se observaron diferencias estadísticamente significativas (p < 0,05), siendo el extracto del líquido intracelular de la cepa L. reuteri NRRL B-14171 el que presentó la mayor capacidad antioxidante (63,77 µME AA), mientras que los que mostraron la menor capacidad antioxidante fueron los fragmentos de pared celular y los ácidos teicoicos de esta misma cepa con 16,14 y 15,99 µME AA, respectivamente.
EVALUACIÓN DE LA CAPACIDAD ANTIOXIDANTE DE L. CASEI SHIROTA Y L. REUTERI NRRL B-14171 EN SACCHAROMYCES CEREVISIAE RC 212
En la figura 2 se presentan los resultados referentes a la evaluación de la capacidad antioxidante de los extractos intra y extracelulares de L. casei Shirota y L. reuteri NRRL B-14171 en Saccharomyces cerevisiae RC 212, lo cual se demostró cuantificando un aumento en las unidades formadoras de colonias por mililitro (UFC/mL) de levadura en comparación con un control positivo expuesto a peróxido de hidrógeno. De acuerdo a los resultados obtenidos, L. casei Shirota mostró un mejor efecto antioxidante al propiciar un mayor crecimiento de Saccharomyces cerevisiae RC 212 en presencia de peróxido de hidrógeno; la bacteria completa, así como los fragmentos de pared celular y sus ácidos teicoicos, mostraron la mejor capacidad antioxidante ya que se contabilizó un crecimiento de 1,065 x 107 UFC/mL, 1,05 x 107 UFC/mL y 1,175 x 107 UFC/mL de Saccharomyces cerevisiae RC 212 en comparación con el control positivo de peróxido, en el cual se contabilizó un crecimiento de 2,04 x 106 UFC/mL de Saccharomyces cerevisiae RC 212; comparado con el tratatamiento negativo, esto representó una reducción del crecimiento del 34, 35 y 27 %, respectivamente. Respecto a L. reuteri ATCC 14171, de forma general, este mostró la menor protección antioxidante frente al peróxido de hidrógeno y únicamente el líquido intracelular mostró la mejor capacidad antioxidante al contabilizarse un crecimiento de 9,4 x 106 UFC/mL de Saccharomyces cerevisiae RC 212 en comparación con el control positivo de peróxido; comparado con el tratatamiento negativo, esto representa una reducción del 42,4 % en el crecimiento de Saccharomyces cerevisiae RC 212. Por otra parte, la bacteria completa, así como los fragmentos de pared celular y sus ácidos teicoicos, mostraron el menor aumento en el crecimiento de Saccharomyces cerevisiae RC 212 al contabilizarse crecimientos de 6,1 x 106 UFC/mL, 6,5 x 106 UFC/mL y 7,9 x 106 UFC/mL, respectivamente.
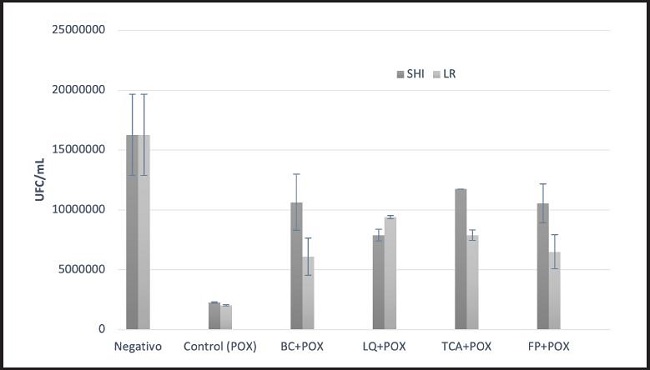
Figura 2. Capacidad antioxidante de extractos intra y extracelulares de L. casei Shirota y L. reuteri en Saccharomyces cerevisiae RC 212 (SHI: L. casei Shirota; LR: L. reuteri; BC: bacteria completa; LQ: líquido intracelular; TCA: ácidos teicoicos; FP: fragmentos de pared celular; POX: peróxido de hidrógeno).
EVALUACIÓN DEL EFECTO ANTIGENOTÓXICO DE L. CASEI SHIROTA Y L. REUTERI NRRL B-14171
La reducción en la cantidad de micronúcleos se muestra en la figura 3. Los extractos intra y extracelulares de ambas cepas evaluadas mostraron capacidad para reducir la genotoxicidad en los linfocitos causada por la acrilamida. En el líquido intracelular de L. reuteri NRRL B-14171 se contabilizaron solo 8,5 núcleos en promedio, comparados con 32,5 registrados en el control tratado con acrilamida, lo cual representa una reducción del 73,8 %; sin embargo, este caso fue estadísticamente similar a la bacteria completa, en la cual se registraron en promedio 11,5 núcleos, lo cual representa una reducción del 64,6 % frente al control. Con respecto a los ácidos teicoicos y la fracción de pared celular, a pesar de que mostraron ser similares estadísticamente, ambos tuvieron un efecto diferente, mostrando una reducción en la formación de micronúcleos de 16 y 14, respectivamente. Por otra parte, el líquido intracelular, los ácidos teicoicos y los fragmentos de pared celular de L. casei Shirota tuvieron un efecto similar en la formación de micronúcleos al contabilizarse 19,5, 18,5 y 20 micronúcleos en promedio, lo cual representa una reducción del 40 %, 43 % y 38,4 %, respectivamente, comparados con el control. Únicamente la bacteria completa de L. reuteri NRRL B-14171 mostró tener una mayor protección sobre la formación de micronúcleos al contabilizarse una reducción del 64,6 % comparada con el control positivo; sin embargo, esta no fue estadísticamente diferente de L. casei Shirota. En la figura 4 se pueden observar los distintos tipos de linfocitos que se observaron en este trabajo, dentro de los cuales aparecen distintas morfologías, como células binucleadas con micronúcleos, células multinucleadas y apoptóticas.
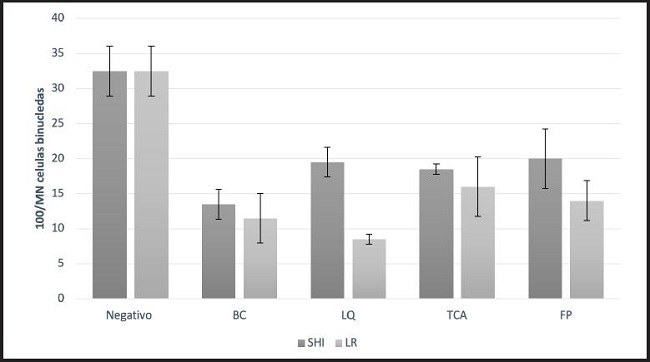
Figura 3. Efecto antigenotóxico de extractos intra y extracelulares de L. casei Shirota y L. reuteri sobre la formación de micronúcleos (MN). SHI: L. casei Shirota, LR: L. reuteri, BC: Bacteria completa, LQ: Líquido intracelular, TCA: Ácidos teicoicos, FP: Fragmentos de pared celular.
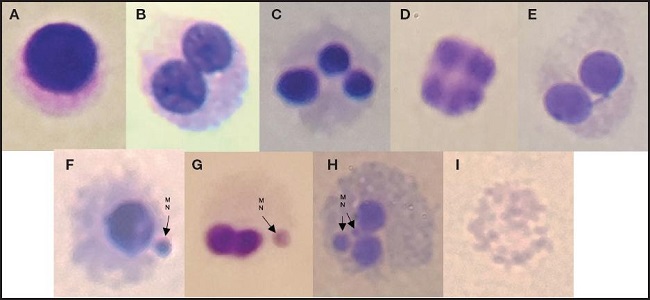
Figura 4. Linfocitos humanos (A: célula normal (mononucleada); B: célula binucleada; C: célula trinucleada; D: célula multinucleada; E: célula binucleada con puente nucleoplasmático; F: célula mononuclear con micronúcleos (MN); G: célula binucleada con 1 MN; H: célula binucleada con 2 MN; I: célula necrótica).
REDUCCIÓN DE LA PEROXIDACIÓN LIPÍDICA EN LINFOCITOS HUMANOS
En la figura 5 se muestra la actividad antioxidante de los extractos intra y extracelulares de L. casei Shirota y L. reuteri NRRL B-14171 en linfocitos humanos. Los extractos intra y extracelulares de L. casei Shirota mostraron tener mayor efecto protector contra la oxidación comparados con los extractos intra y extracelulares de L. reuteri NRRL B-14171. La bacteria completa, así como los ácidos teicoicos de L. casei Shirota, mostró la mayor protección contra la peroxidación lipídica en los linfocitos ya que, en el caso de la bacteria completa, se registraron 0,0004 mMolTEP y en el caso de los ácidos teicoicos 0 mMolTEP, lo cual representa una reducción de la peroxidación de la membrana del eritrocito del 85 y el 100 %m respectivamente para estos extractos comparados con el control. Por otra parte, el líquido intracelular de esta cepa también mostró ser uno de los extractos que más protección mostró ante la peroxidación lipídica, al contabilizarse 0,00057 mMolTEP (80 % menos que el control). Respecto a L. reuteri NRRL B-14171, los ácidos teicoicos fueron los únicos extractos que mostraron la mayor reducción de la peroxidación lipídica, al contabilizarse 0,0008 mMolTEP, lo que representa un 70 % de reducción comparada con el control. Por su parte, los ácidos teicoicos de L. casei Shirota mostraron el mayor efecto antioxidante, siendo estos significativamente diferentes a los extractos de la pared celular, líquido intracelular y bacteria completa. En el caso de la fracción de pared celular, esta mostró mayor efecto antioxidante que la misma bacteria completa, lo cual podría deberse a que, al estar fraccionada, presenta grupos funcionales o cargas que podrían estar influenciando el efecto antioxidante. Por otra parte, con respecto a L. reuteri NRRL B-14171, el extracto intracelular correspondiente al líquido intracelular fue el único que mostró un mayor efecto antioxidante, siendo estadísticamente diferente a los otros extractos. La pared celular presentó un efecto antioxidante similar estadísticamente al de la bacteria completa y los ácidos teicoicos al fragmento de la pared celular.
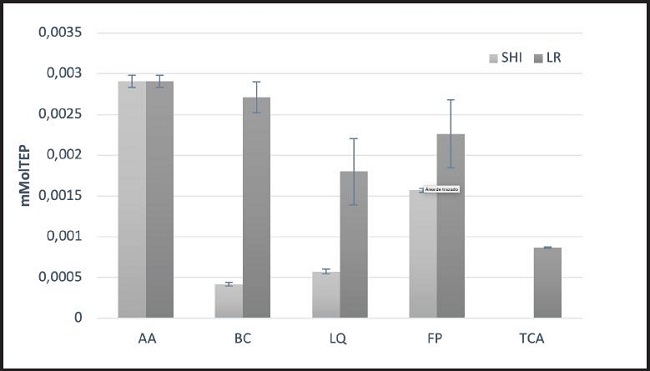
Figura 5. Reducción de la peroxidación lipídica en linfocitos humanos por los extractos intra y extracelulares de L. casei Shirota y L. reuteri (SHI: L. casei Shirota; LR: L. reuteri; AA: acrilamida; BC: bacteria completa; LQ: líquido intracelular; TCA: ácidos teicoicos; FP: fragmentos de pared celular).
DISCUSIÓN
De acuerdo con nuestros resultados, las bacterias acidolácticas evaluadas mostraron que eran capaces de proteger contra la oxidación ocasionada por la acrilamida, variando la capacidad antioxidante significativamente en los extractos intra y extracelulares y mostrando una marcada diferencia entre las dos cepas evaluadas, lo cual indica que esta actividad depende de cada cepa. De acuerdo a nuestros resultados, los extractos intracelulares son los que mostraron mayor actividad antioxidante y, de estos, el extracto intracelular de Lactobacillus reuteri NRRL B-14171 fue el que presentó mayor capacidad antioxidante (63,77 µMolar de equivalentes de ácido ascórbico), lo cual representa un 45 % más que la bacteria completa, la pared celular y los ácidos teicoicos. Estos resultados son similares a lo reportado por Cuevas-González y cols. (9), que reportaron una alta capacidad antioxidante para el líquido intracelular de las bacterias acidolácticas evaluadas. Adicionalmente, Aguilar-Toala y cols. (10) reportaron que el líquido intracelular de trece cepas de lactobacilos mostró una mayor actividad antioxidante que la pared celular y que la bacteria completa y, de estas, Lactobacillus casei CRL431 presentó la mayor capacidad antioxidante seguida de las cepas Lactobacillus fermentum B1932 y Lactobacillus casei DCP3968. Estos resultados demuestran que ciertas cepas de lactobacilos podrían ser una buena fuente de posbióticos y una alternativa diferente como protección contra la oxidación causada por compuestos tóxicos adquiridos a través de la dieta.
Diversos autores indican que la actividad antioxidante de algunas cepas de lactobacilos podría atribuirse a la producción de compuestos ubicados en la superficie celular, como polisacáridos extracelulares, ácidos teicoicos y lipoteicoicos (11-13). Con respecto a la actividad antioxidante de los extractos intra y extracelulares de L. casei Shirota y L. reuteri NRRL B-14171 en los linfocitos humanos, los extractos intra y extracelulares de L. casei Shirota mostraron tener mayor efecto protector contra la oxidación comparados con los extractos intra y extracelulares de L. reuteri NRRL B-14171. Los ácidos teicoicos de L. casei Shirota mostraron mayor efecto antioxidante, siendo estos significativamente diferentes a los extractos de la pared celular, líquido intracelular y bacteria completa. En el caso de la fracción de pared celular, esta mostró mayor efecto antioxidante que la misma bacteria completa, lo cual podría deberse a que, al estar fraccionada, presenta grupos funcionales o cargas expuestos que podrían estar influenciando el efecto antioxidante. Por otra parte, con respecto a L. reuteri NRRL B-14171, el extracto intracelular correspondiente al líquido intracelular fue el único que mostró un mayor efecto antioxidante, siendo estadísticamente diferente a los otros extractos; de ellos, la pared celular presentó un efecto antioxidante estadísticamente similar al de la bacteria completa y los ácidos teicoicos al del fragmento de la pared celular.
El uso de levaduras como modelo biológico para el estudio de antioxidantes es un modelo simple pero confiable que puede emplearse para evaluar la protección contra la oxidación a través de la reducción del crecimiento de una célula, además de que varias cepas de lactobacilos han demostrado ser resistentes al peróxido de hidrógeno a diferentes concentraciones (14,15). Al respecto de la capacidad antioxidante de los extractos intra y extracelulares de L. casei Shirota y L. reuteri NRRL B-14171 en Saccharomyces cerevisiae RC 212, L. casei Shirota mostró un mejor efecto antioxidante al propiciar un mayor crecimiento de Saccharomyces cerevisiae RC 212 en presencia de peróxido de hidrógeno. La bacteria completa, así como los fragmentos de pared celular y sus ácidos teicoicos, mostraron la mejor capacidad antioxidante al permitir solo una reducción del crecimiento de Saccharomyces cerevisiae RC 212 del 34, 35 y 27 % para la bacteria completa, los fragmentos de pared celular y los ácidos teicoicos, respectivamente. Estos resultados coinciden con los resultados hallados sobre la capacidad antioxidante de los extractos obtenidos, en donde la pared celular y los ácidos teicoicos de esta cepa fueron los que mostraron la mejor capacidad antioxidante. Adicionalmente, L. reuteri NRRL B-14171 mostró la menor protección antioxidante frente al peróxido de hidrógeno y únicamente el líquido intracelular mostró la mejor capacidad antioxidante al permitir la reducción del 42,4 % del crecimiento de Saccharomyces cerevisiae RC 212. Este resultado es consistente con los encontrados respecto a la capacidad antioxidante y la protección contra la oxidación evaluada en los linfocitos ya que, en ambos casos, el líquido intracelular de la misma cepa mostró la mejor capacidad antioxidante y protección contra la oxidación.
Respecto a la evaluación del efecto antigenotóxico de los extractos intra y extracelulares mediante la formación de micronúcleos, se observó que todos los extractos analizados presentaron efecto protector contra la genotoxicidad causada por la acrilamida. El análisis estadístico reveló que no hubo diferencias significativas entre los diferentes extractos extra e intracelulares y la bacteria completa con respecto a la cantidad de micronúcleos formados respecto al grupo negativo; sin embargo, el líquido intracelular de L. reuteri NRRL B-14171 presentó la menor formación de estructuras binucleadas con un promedio de 8,5 células, correspondiente a una reducción del 73,8 % respecto a las 32,5 células del grupo adicionado solo con acrilamida, lo que lo distingue como el extracto con mayor efecto antigenotóxico, mientras que el de menor protección corresponde al extracto de fragmentos de pared de L. casei Shirota, con un 38,4 % de inhibición en la formación de micronúcleos (20 MN). La acrilamida se ha visto involucrada en el desarrollo de múltiples afectaciones comprobadas a partir de análisis tanto in vivo como in vitro, en donde el daño al material genético es evidente; por ello se buscan alternativas que ayuden a atenuar dichos efectos, tal como lo hicieron Zamani y cols. (8), quienes, de forma similar a nuestra investigación, observaron una disminución de la formación de células binucleadas en los linfocitos de sangre humana, utilizando L-carnitina como agente antioxidante. Salimi y cols. (16), por su parte, utilizaron el ensayo de 8-hidroxi-2’-desoxiguanosina (8-OHdG) para evaluar la disminución del daño al ADN utilizando ácido elágico, observando una disminución significativa en la formación de aductos con 8-OHdG al utilizar concentraciones de 25 y 50 µM del ácido. Por otro lado, se cuenta con poca información sobre la capacidad que tienen los componentes extra e intracelulares de los lactobacilos para contrarrestar la genotoxicidad causada por la acrilamida y solo se ha comprobado que el uso de Lactobacillus y de componentes como los ácidos teicoicos puede generar una importante fijación con la acrilamida, favoreciendo su degradación y eliminación (17,18), lo que podría ser un factor clave en la disminución del daño genético.
Estudios previos han indicado que la acrilamida induce la formación de especies reactivas de oxígeno en los linfocitos humanos. La generación de estas es inducida por la acrilamida, pudiendo afectar a macromoléculas importantes, como son las proteínas y lípidos de membrana, e inducir la peroxidación de los lípidos en las células (16). En el presente trabajo, los ácidos teicoicos de ambas cepas evaluadas mostraron la mayor actividad antioxidante cuando se expusieron los linfocitos a la acrilamida. La bacteria completa y los ácidos teicoicos de L. casei Shirota mostraron la mayor protección contra la peroxidación lipídica en los linfocitos, al contabilizarse una reducción del 85 y el 100 %, respectivamente, para estos extractos. Por otra parte, el líquido intracelular de esta cepa también mostró ser uno de los extractos de mayor protección ante la peroxidación lipídica, al contabilizarse una reducción del 80 %. Respecto a L. reuteri NRRL B-14171, los ácidos teicoicos fueron los únicos extractos que mostraron una mayor reducción de la peroxidación lipídica, al contabilizarse un 70 % de reducción.
CONCLUSIONES
Ambas cepas evaluadas, así como sus extractos intra y extracelulares, mostraron tener capacidad de contrarrestar el estrés oxidativo y la genotoxicidad causados por la acrilamida. Los fragmentos de pared celular de L. casei Shirota y L. reuteri NRRL B-14171, así como el líquido intracelular de esta última, mostraron la mayor capacidad antioxidante. Sin embargo, se encontraron marcadas diferencias respecto a esta propiedad en ambas cepas. Estas diferencias observadas podrían estar determinadas por la variación de la composición estructural de los ácidos teicoicos y lipoteicoicos, además de la conformación de los peptidoglicanos y de elementos presentes en el líquido intracelular que podrían tener mejor repuesta al liberarse tras el rompimiento celular y que podrían neutralizar los radicales producidos por la acrilamida mediante la liberación de agentes reductores.
Los resultados encontrados sugieren que el empleo de extractos intra y extracelulares de Lactobacillus reuteri NRRL B-14171 y Lactobacillus casei Shirota podría ser una alternativa para reducir los efectos de genotoxicidad y estrés oxidativo causados por la acrilamida, sin necesidad de requerir una estructura viable sin alteración de su estructura.

