










 Servicios personalizados
Servicios personalizados
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroduction
Most of research on the acquisition of simple discriminations is based on procedures in which a single stimulus works as the positive stimulus (S+), another one works as the negative stimulus (S-), and a single type of reinforcer stimulus is used (De Rose, McIlvane, Dube, Galpin, & Stoddard, 1988). Commonly, if the animal emits the target response in the presence of the S+, the reinforcer stimulus (usually food) will appear, while the same response will be punished or extinguished when it is given in the presence of the S- (Polín & Pérez, 2017). This is what we call “the three-term contingency” (Skinner, 1938; 1953; 1969), or, in other words, the simplest possible case of interrelationship between the antecedent events, the responses, and the consequences in operant conditioning.
However, it could be likely that in uncontrolled environments (outside the laboratory) the number of stimuli which are involved in the acquisition of these kinds of discriminations would increase. Therefore, it would be reasonable to think that the most widely used procedures do not conform to these phenomena as much as they could. Hence, the present study aimed to address this question by evaluating if a simple discrimination is acquired faster as a function of the variety (or not) of negative stimuli.
In uncontrolled environments, certain responses that fall under excitatory control of a S+ can also be exposed to the inhibitory control of several S- (and not exclusively a single one). For example, the response “to say table" can be reinforced in the presence of a table (S+), but the number of events in whose presence “saying table" is not reinforced (or is even punished) is not necessarily limited to one. So, the tact (Skinner, 1957) “table" would be extinguished (or punished) in the presence of a chair, a bed, a sofa, or, in short, anything other than a table.
In this sense, it is remarkable that several authors have shown interest in studying the possible effect of varying the events which compose the contingencies (that is to say, not using always the same ones) on different learning processes. The variety of events has been applied both to the antecedent (Farthing, 1974; Mandler, 1970, 1973; Mullins & Winefield, 1979; Pérez & Polín, 2016; Schaeffer & Shandro, 1969; Walk & Saltz, 1965; Williams, 1967, 1968) and the consequent (Bowman, Piazza, Fisher, Hagopian, & Kogan, 1997; Egel, 1980, 1981; Milo, Mace, & Nevin, 2010; (Polín & Pérez, 2017; Steinman, 1968a, 1968b; Thrailkill, Epstein, & Bouton, 2015).
Regarding the specific case of the antecedent stimuli, the experimental evidence suggests that different results should be expected as long as the amount of stimuli is increased. For example, two experiments conducted by Williams (1967; 1968) showed that pigeons acquire a color simultaneous discrimination faster with seven, rather than two, negative stimuli. This time, the author explained the results as a function of the greater aversion generated by the larger number of S-.
A different phenomenon was reported in another two experiments which involved simultaneous discriminations in a Y maze with rats (Mandler, 1970), although in this case the number of positive stimuli was also manipulated, resulting in two different conditions: constant S+ with multiple S-, and constant S- with multiple S+. The results showed that the discrimination was easier to acquire for the animals of the constant S- and multiple S+ condition. The same author also observed a differential effect due to the variety of stimuli (again, both S+ and S-) on transfer tests (Mandler, 1973). Specifically, transfer was effective on the basis of the constant stimulus, either it had previously been positive or negative. We can also find another study which showed a worse acquisition of a simultaneous discrimination in rats with varied negative stimuli than with varied positive stimuli (Mullins & Winefield, 1979).
Nevertheless, there is also a study conducted with rats in which the authors found no differences in the acquisition of a simultaneous discrimination as a function of the number of negative stimuli (Schaeffer & Shandro, 1969). These authors pointed out that this lack of differences could have been due to a very quick acquisition, and, in fact, it has been generally shown that it is easier to acquire a simultaneous, rather than a successive, discrimination (Saunders & Green, 1999). However, they observed that a reversal task was systematically and consistently better performed by the animals who had been trained with the larger number of S-.
Hence, the available evidence so far suggests two different conclusions: On one hand, the fact that applying a variety of S- affects the results of simple discrimination training (although the way in which they are affected is not so clear). On the other hand, it seems that these different phenomena are strongly sensitive to certain specific procedural manipulations.
Experiment 1
Continuing with the example given at the beginning of this manuscript, outside the laboratory, the response “table” would be reinforced in the presence of a table and would be punished or extinguished in the presence of anything other than a table. And, in turn, those stimuli which are not a table (a chair, a bed, a sofa…), would acquire S+ functions for their appropriate responses (“chair”, “bed” or “sofa”), as well as S- functions for the response “table”. In this example, the chair, the bed and the sofa are different stimuli which have one important feature in common: they are not a table. So, all of them could form a functional set (Vaughan, 1988) and be taught as discriminative stimuli for the response “not a table”.
In the present experiment, pigeons were trained in a “go/no-go” simple discrimination which provided reinforcement for both “go” (peck) and “no go” (no peck) responses, depending on the stimulus which had been presented (S+ and S-, respectively).
The aim of the experiment was to check whether reinforcing “no go” responses in the presence of a wide variety of stimuli could lead (or not) to a faster acquisition compared to reinforcing these responses in the presence of a constant stimulus.
Methods
Participants
Five rock pigeons (Columba livia) were kept at approximately 85% of their Free-Feeding Weight (FFW). All of them had previously been used for the same experiment on choice behavior (which did not keep any kind of procedural relationship with the one that is here described). Water and pigeon grit were always available in their home cages. The temperature and humidity of the room was maintained stable and a 12:12 hour light-dark cycle was employed during the whole experiment (the lights turned on at 10 am).
Instruments
Four modified operant chambers for bird conditioning with a tactile PC screen attached on the left side were used. Each chamber was 43.5 cm high, 64 cm long and 45 cm wide. The front of the chamber was equipped with a feeder providing a mix of grain and with three standard keys (left, central, right). In the middle of the back side, a 35 watt white light provided illumination. The left panel of the chamber was removed, and a touch screen monitor was installed instead (ELO Touchsystems MODEL ETL 121-C-75WB-1). The monitor displayed a resolution of 800 per 600 pixels, 60Hz, SVGA (16.2 million of colors). The modified operant chambers were enclosed in sound-attenuating hulls. A ventilation fan installed in each hull produced a white noise, masking extraneous sounds. Each touch screen was connected to an IBM PC compatible. A tailor made program (DV) controlled the presentation of stimuli and recorded the responses. Each PC was connected to a MED R/M interface cabinet SG - 6001C SN controlled by an IBM compatible computer. The software used to run the experiments, to control the presentation of the stimuli, and to record the responses was MedPC 2.0 for Windows.
The stimuli appeared through the left key and through the screen. The antecedent stimuli (S+ and S-) were presented through the touch screen monitor (only one at the same time), while the responses were always registered in the white illuminated left key (the nearest to the screen). The antecedent stimuli consisted of the whole screen illuminated in different colors (one color for each stimulus). The colors wavelengths were the following: 440, 450, 460, 480, 500, 520, 540, 560, 580, 600, 620, 640, and 750 nm.
Four seconds of access to food (grain) was always used as the reinforcer stimulus.
Procedure
Before the beginning of the experiment, all the pigeons were auto-shaped until a white left key-pecking response was established at a consistent level. Specifically, the criterion to finish this phase was to key-peck the 90% of trials in which the key was illuminated.
Acquisition of the two simple discriminations
The subjects were divided into two conditions: varied and constant.
A Fixed Ratio (FR) 3 schedule was designed, using a “go/no-go” successive simple discrimination procedure in which the S+ and the S- appeared an average of 50% of the trials each. For both conditions, sessions consisted of 40 intermixed trials, therefore, S+ and S- were presented an average of 20 times each per session with a 45 second inter-trial interval (ITI).
Both S+ and S- remained activated until the subjects responded (FR 3) or until 10 seconds passed. We chose to require white left key-pecking rather than pecking directly to the S+ in order to isolate operant and conditioned responses, so, when the S+ or the S- appeared, the left key was illuminated until the response (FR 3) was given or until 10 seconds passed.
Trials in which subjects responded pecking to the white left key in the presence of the S+, as well as trials in which they did not respond in the presence of the S-, were considered correct (reinforced with food). Those trials in which subjects responded pecking to the left key in the presence of the S-, and those trials in which they did not respond in the presence of the S+, were considered failures (not reinforced).
The number of sessions needed to reach the acquisition criterion (90% of success trials in a single session) was measured as a dependent variable. When this criterion was reached, the discrimination was extinguished for all the subjects (same procedure as in the acquisition, but, this time the food never appeared regardless of the stimulus that had been presented and regardless of the behavior of the subjects) until they completely stopped responding, and then, they were all again auto-shaped before being exposed to a new discrimination, with a new stimuli set, changing the condition. Therefore, subjects that were assigned to the varied condition in discrimination 1, changed to the constant condition in discrimination 2. Subjects that were assigned to the constant condition in discrimination 1, changed to the varied condition in discrimination 2.
In the varied condition, the S- stimulus could be one out of seven different colors, varying randomly from trial to trial. In the constant condition, the S- stimulus was always the same, being its value (measured in nm) the average of all the stimuli used in the varied condition.
In the first discrimination, the S+ consisted of the whole screen illuminated in a color of 450 nm for both conditions. For the varied condition, the S- consisted of the whole screen illuminated in a color which varied randomly across the following wavelengths: 520, 540, 560, 580, 600, 620, or 640 nm. For the constant condition the S- was always 580 nm.
In the second discrimination, the S+ was 750 nm for both conditions. For the varied condition, the S- could randomly be 440, 460, 480, 500, 520, 540, or 560 nm. For the constant condition, the S- was always 500 nm.
Pigeon 1 (P1) and pigeon 3 (P3) were first exposed to the varied condition and then to the constant. Pigeon 4 (P4), pigeon 5 (P5) and pigeon 6 (P6) were first exposed to the constant condition and then to the varied.
Results and discussion
Regardless of the condition to which each subject was first assigned, there was a differential effect which can be directly observed taking a look to the individual subject data. Specifically, three out of five pigeons acquired the discrimination faster with constant, rather than varied, negative stimuli. Figure 1 depicts these within-subject comparisons.

Figure 1 Acquisition curves: individual subject data regarding the percentage of correct trials along sessions in both simple discriminations as a function of the condition (V, varied; C, constant).
It can be observed that P3 and P5 acquired the two discriminations very fast and with no speed differences between them. P3 just needed two sessions in both conditions and P5 needed two for the varied condition and three for the constant. Nevertheless, the results regarding P1, P4, and P6 clearly show that the discrimination was easier to acquire in the constant condition than in the varied one. For the constant condition, P1, P4, and P6 reached the acquisition criterion in four, six, and three sessions, respectively. For the varied condition they needed 10, 12, and 19, respectively.
Taking into account the two discriminations, the mean number of sessions needed to reach the acquisition criterion was 9 for the varied condition and 3.6 for the constant.
Regarding the extinction phases, the results were the following: P1 needed six sessions for the first discrimination and nine for the second. P3 needed five for the first one and seven for the second. P4 needed five sessions for both discriminations. P5 needed five for the first one and four for the second. P6 needed three sessions for both discriminations.
The mean number of sessions needed to reach the extinction criterion was 4.6 for the varied condition and 5.8 for the constant. Hence, the difference between the two conditions was too slight to be considered relevant.
The results show that a variety of S- which also function as S+ for other responses different than key-pecking could lead to a slower acquisition compared to the same type of training involving just a single S-. Actually, there was something that the subjects could do in order to obtain food in the presence of the S-. So, we could argue that two different responses were being explicitly trained: key-pecking and not key-pecking. Accordingly, it would be reasonable to think that if a “not key-pecking” response had to be acquired in the presence of several stimuli, the process would take longer than if this response had to be controlled only by one stimulus.
A go/no-go procedure with such features could also result in a phenomenon closely related to the ‘superstition’ (Skinner, 1948): In the reinforced “no-go” trials there was a contingency relationship between the “waiting behavior” and the food presentation (and this relationship did not exist in Skinner´s experiment), but it could be possible that each pigeon consistently responded (differently from each other) in the same way every time that a S- was presented. Indeed, in our procedure, every response (different from key-pecking) was contingently reinforced.
Experiment 2
In the previous experiment, reinforcing “no go” trials resulted in two different responses that were explicitly being trained. Therefore, in Experiment 2 we decided to use a standard go/no-go procedure, so that when the S- appeared (“no-go” trials), waiting for ten seconds was not reinforced. In this manner, the different S- never correlated with food and every response given in their presence was extinguished.
This procedure is also compatible with the formation of a functional set (Vaughan, 1988), but, this time the set of S- should only acquire one function (S-).
The main rationale for carrying out this experiment was to check if the previously observed variety effect could also arise without reinforcing a second response. This procedural manipulation could allow us to assess the effect of the inclusion of a variety of S- without these stimuli acquiring more functions than those of S-. And, more specifically, without acquiring excitatory control over an explicitly trained response.
Methods
Participants
Six rock pigeons (Columba livia) were kept at approximately 85% of their FFW. All of them had previously been used for the same experiment on choice behavior (the same one as the pigeons of Experiment 1). Water and pigeon grit were always available in their home cages. The temperature and humidity of the room was maintained stable and a 12:12 hour light-dark cycle was employed during the whole experiment (the lights turned on at 10 am).
Instruments
The operant chambers, touch screen monitors, software, and sets of stimuli were the same as those used in Experiment 1.
Procedure
The two conditions were the same as shown in Experiment 1 (varied and constant). Pigeon 7 (P7), pigeon 8 (P8) and pigeon 11 (P11) were first exposed to the varied condition and then to the constant. Pigeon 9 (P9), pigeon 10 (P10) and pigeon 12 (P12) were first exposed to the constant condition and then to the varied. The pigeons were initially auto-shaped before the beginning of the acquisition phase and again before starting the second discrimination training (after being exposed to the first discrimination extinction).
Regarding the FR 3 schedule, the number of trials, the ITI duration, the success criterion, and the rest of features of the procedure, all the sessions were conducted in the same way as described in Experiment 1.
There was only one procedural difference between this experiment and the first one: in Experiment 2, waiting for 10 seconds in the presence of the S- was not reinforced with food (although it was considered a correct trial).
Results and discussion
As shown in the individual subject data (Figure 2), four out of six pigeons acquired the discrimination faster with constant, rather than varied, negative stimuli. The two left pigeons acquired the discrimination faster with varied, rather than constant, negative stimuli.
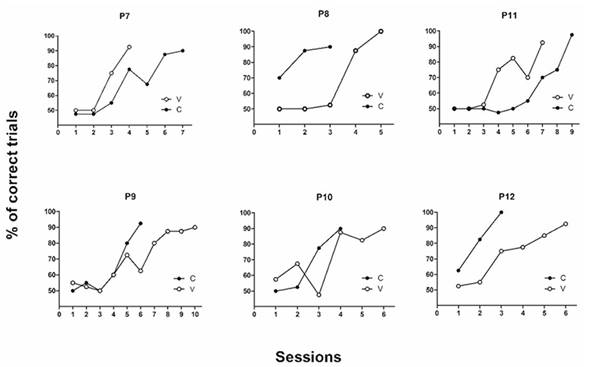
Figure 2 Acquisition curves: individual subject data regarding the percentage of correct trials along sessions in both simple discriminations as a function of the condition (V, varied; C, constant).
P7 and P11 needed more sessions to acquire the discrimination when they were exposed to the constant condition (seven and nine sessions, respectively) than when they were exposed to the varied (four and seven sessions, respectively). On the contrary, P8, P9, P10 and P12 needed more sessions to complete the varied condition (5, 10, 6 and 6 sessions, respectively) than to complete the constant (3, 6, 4 and 3, respectively).
Furthermore, regardless of the condition (“varied-constant” versus “constant-varied”), all the pigeons except P8 acquired their first discrimination faster than their second one.
Taking into account the two discriminations, the mean number of sessions needed to reach the acquisition criterion was 6.3 for the varied condition and 5.3 for the constant.
Therefore, although four out of six pigeons acquired the discrimination faster with constant, rather than varied, negative stimuli, the mean number of sessions needed to reach the acquisition criterion was very similar in both conditions. There was a slight difference between them, but it was not as clear as observed in Experiment 1.
As described above, there is a fact in our results that may contribute to explain this lack of overall differences: Five of the six pigeons acquired the first discrimination faster than the second one, regardless of the condition to which they were first assigned. These data suggest that a presentation order effect might have occurred. Hence, we could think that the similarity between the stimuli used for discrimination 1 and for discrimination 2 could probably be at the basis of these results. But the stimuli that we used were the same as those described in Experiment 1 and this effect was not observed before, so, we have to focus on the procedural differences between both experiments in order to explain why it happened this time.
Regarding the extinction phases, the results were the following: P7 needed four sessions for the first discrimination and six for the second. P8 needed three for the first one and six for the second. P11 needed two sessions for the first and four for the second. P9 needed five sessions for the first and four for the second. P10 needed five for the first and seven for the second. P12 needed four and nine, respectively.
The mean number of sessions needed to reach the extinction criterion was 4.8 for the varied condition and 5 for the constant. Therefore, there were no differences between the two conditions with regard to the resistance to extinction.
General discussion
On one hand, results regarding Experiment 1 suggest that it is easier to acquire a simple discrimination with constant, rather than varied, negative stimuli (using a go/no-go procedure in which “no-go” trials are also reinforced).
These results do not support those reported in previous studies (Schaeffer & Shandro, 1969; Williams, 1967, for example), although there is an important procedural difference between all of them and the present experiment which could probably explain why the results are different from each other: In the cited studies, the tasks consisted of different simultaneous discriminations in which both the S+ and the S- were presented at the same time. Furthermore, these authors increased the number of the same S-, so, we cannot talk about varied or different stimuli. In our experiments, the varied condition meant that every time that the S- was presented, it could be different from the previous one, varying randomly across seven different stimuli.
This difference has an important implication when analyzing and explaining the results. For example, Williams (1967) reported that the performance of the rats was better with seven S- presented at the same time than with two S- and explained the effect in terms of a greater aversiveness regarding the larger number of S- because of the larger number of non-rewarded responses given in their presence. In fact, the probability of success if the animals responded randomly was one out of eight in the seven S- condition and one out of three in the two S- condition.
In our first experiment, the probability of success was controlled (always 50%, regardless of the condition and regardless of the presence of S+ or S-), so, if we assume the hypothesis of the aversiveness, our findings do not contradict Williams´, but provide evidence that, when this variable is controlled, they must be explained in other terms: It seems that the variety of negative stimuli in a successive discrimination could add more complexity to the context, which leads to a lower acquisition rate.
More specifically, this complexity could be determined by the lower number of trials with each S- compared to the constant condition. Indeed, the number of trials was seven times lower in the varied condition than in the constant one, so, the latter provided more amount of training per session with the single S- than the former.
On the other hand, results regarding Experiment 2 show that the phenomenon can only be observed when the animals had first been exposed to the constant condition. These results suggest that a negative transfer effect between the two discriminations can be observed (with the sets of visual stimuli that we have used).
In effect, although the value of the wavelengths was a controlled variable, some of them looked very similar to each other. For example, the color which functioned as S+ in the first discrimination was blue (450 nm), and so they were some of the colors which functioned as S- in the second discrimination (440, 460, 480, and 500 nm). The same happened with some of the colors which functioned as S- in the first discrimination (580, 600, 620, and 640 nm) and the one which functioned as S+ in the second one (750 nm): they all looked red or very similar to red. Hence, the pigeons should have initially made more failures in the second discrimination for this reason, which would explain the results (that is to say, acquisition should have been slower in the second discrimination than in the first one).
However, we used the same sets of stimuli in both experiments, so, why was the negative transfer effect only observed in the second one? Maybe a functional analysis based on the procedural differences between the experiments can answer this question: In Experiment 1, the reinforcer stimulus could also appear in “no-go” trials if the pigeons waited for 10 seconds, while in Experiment 2, every response given during “no-go” trials was extinguished. So, in Experiment 1 all the S- (for both conditions) were also functioning as S+ for other responses different from pecking the key. This means that the appearance of the S- could also correlate with food, facilitating the responses in its presence (or in the presence of a very similar stimuli) in the second discrimination and preventing, in this manner, the negative transfer effect.
Considering the results of both experiments, it could be argued that the stimuli which functioned as S- formed a functional class. Generally, it is assumed that several stimuli which signal the appearance of the same consequence (reinforcing or not) could be considered functionally equivalent (Dube, McIlvane, Maguire, Mackay, & Stoddard, 1989; Zentall, Wasserman, & Urcuioli, 2014). Therefore, a delay in acquisition should be expected under the conditions that require the formation of a functional class (compared to the conditions which only include a constant stimulus).
Further research is needed in order to keep on deepening on the basic features of the phenomenon, as well as on its possible implications of applied nature. But, globally taken, our results provide evidence that a simple discrimination is differentially acquired as a function of the variety (or not) of the antecedent stimuli involved in the process. Likewise, they support previous findings that suggest that this effect is sensitive to certain procedural features and, furthermore, two of them have been identified: the sensory modality of the stimuli and the type of go/no-go procedure.
These conclusions could be useful for behavior analysis, emphasizing in the importance of including the variety of antecedent stimuli as a relevant variable to consider when difficulties in learning do arise, either in clinical or educational contexts, and even for non-human animals training.
Given the results of our experiments, it does not seem recommendable to include a variety of negative stimuli at the beginning of training, as it could lead to a slower acquisition.

