










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Española de Enfermedades Digestivas
versión impresa ISSN 1130-0108
Rev. esp. enferm. dig. vol.97 no.7 Madrid jul. 2005
| ORIGINAL PAPERS |
Antisense gene therapy using anti-k-ras and antitelomerase oligonucleotides
in colorectal cancer
S. Lledó, R. Alfonso and S. F. Aliño1
Department of General and Digestive Surgery. Hospital Clínico Universitario. Departments of Surgery and 1Pharmacology.
Universidad de Valencia. Valencia, Spain
ABSTRACT
Aim: to test the efficacy of anti-k-ras and antitelomerase oligonucleotides for disabling colorectal cancer cell growth.
Material and methods: an established human colorectal cancer cell line (SW 480, ATTC®) was used. Oligodeoxiribonucleotides (ODNs) have a phosphorotioate modification to ensure intracellular intake. We used an antitelomerase ODN (Telp5) and two anti-k-ras ODNs (AS-KRAS and ISIS). AS-KRAS is designed to join the k-ras oncogene's exon 1. ISIS links to the terminal transcription unit 5' of k-ras. Telp5 joins the template region of the hTR telomerase subunit. ODNs have been tested in different concentrations (1, 5, 10, 20 micromolar). Cell viability has been tested at 48 and 72 hours. Statistical analysis and graphic design were made with the statistical package "Analyzing Data with GraphPad Prism-1999", GraphPad Sofware Inc., San Diego CA©. We used the Student's t test for statistical analysis.
Results: the lowest dose (1 µM) was not effective. Using the highest dose (20 mM for 48 hours) of combined AS-KRAS and Telp5 cell viability decreased to 99.67%. The rest of results varied depending on ODN type, dose, and exposure time.
Conclusions: tested antisense ODNs stop colorectal cancer cell growth, and a combination of anti-telomerase and anti-k-ras is the most useful treatment. Efficacy is best with a higher dose and longer treatment period.
Key words: Colorectal cancer. Oligoribonucleotides. K-ras oncogene. Telomerase. Antisense therapy.
Lledó S, Alfonso R, Aliño SF. Antisense gene therapy using anti-k-ras and antitelomerase oligonucleotides in colorectal cancer. Rev Esp Enferm Dig 2005; 97: 472-480.
Recibido: 13-09-04.
Aceptado: 04-01-05.
Correspondencia: Salvador Lledó Matoses. Servicio de Cirugía-5º C. Hospital Clínico Universitario. Avda. Blasco Ibáñez, 17. 46010 Valencia. e-mail: cirugia@telefonica.net
INTRODUCTION
Colorectal cancer (CRC) survival after curative surgery depends on local and distant recurrence. After curative resection 5-year survival varies from 40 to 80% depending on local recurrence rates (1). With a 100% control of local disease, 17% of patients will die from cancer; with a local control of 80%, 31% of deaths will occur (2). These recurrences may result from tumoral cell spread during surgery (3,4).
The K-ras oncogene is activated in almost 70% of human CRCs (5). This is partly responsible for the uncontrolled growth of CRC cells (6). Its activation is also related to vascular endothelial growth factor (VEGF) hyperexpression, which is involved in angiogenesis (7).
DNA in telomeres shortens with each cellular division. In normal cells DNA shortens with each DNA replication, and therefore chromosomes become unstable, and cells cannot divide and eventually die.
Nevertheless, more than 90% of cancer cells have increased telomerase activity, which contributes to tumor cell uncontrolled growth (8). Telomerase activity is increased in 90-100% of human CRCs (9,10). This fact has been proposed as an alternative tumor development pathway (11).
Antisense techniques for gene therapy consist in blocking the expression of a gene using an oligonucleotide that is complementary to the gene sequence whose blocking is desired (12). Such an antisense nucleotide can be chemically designed, the simplest way to do so being the generation of a small piece of DNA: an oligodeoxiribonucleotide. Antisense sequences are designed in such a way that they recognize and link specific mRNA sequences. In this way, the double-chain hybrids generated cannot be translated by the ribosome into a protein, and protein translation cannot take place.
In our study antisense oligodeoxiribonucleotides (AS-ODN) have been designed to interfere with K-ras and telomerase (hTERT subunit) genes. The aim of this study was to evaluate growth inhibition achieved in vitro in an established CRC cell line.
MATERIAL AND METHODS
An established human cell line was used: SW 480 (ATCC. The Global Bioresource Center®). This cell line comes from primary colorectal adenocarcinomas in stage B of Dukes' classification. It has a mutation in codon 273 of the p53 gene and a mutation in codon 12 of the protooncogen k-ras.
Cell viability studies have been made with the commercial kit Calcein-AM (Molecular Probes®). This procedure allows to identify living cells after incubation with modified calcein. Calcein-AM has a fluorescent activity that develops only when metabolized by viable cells.
AS-ODNs used have a phosphorotioate modification in order to prevent degradation by cellular nucleases, so intracellular intake is ensured. We designed an antitelomerase AS-ODN (Telp5) and two anti k-ras AS-ODNs (AS-KRAS and ISIS). The sequences of each AS-ODN are: ISIS 6957: 5'-CAG TGC CTG CGC CGC GCT CG-3'; AS-KRAS: 5'-AAG TTT ATA TTC AGT CAT-3'; Telp5: 5'-CCC TTC TCA GTT AGG GTT AG-3'.
ISIS 6957 has been previously used by other groups (13,14). The AS-KRAS tested in our study was designed to link exon 1 of the k-ras oncogene. ISIS joins the 5' terminal transcription unit of the k-ras oncogene. Telp5 links the telomerase hTR subunit. AS-ODNs were tested in different concentrations: 1, 5, 10 and 20 µM. We selected these amounts based on studies by other groups (13,15,16), but taking into consideration that we used no transfection vectors to facilitate cellular intake. No cytotoxicity has been described in vitro with concentrations under 100 µM, nor toxicity in murine or ape models using intravenous doses under 100 mg/kg (12).
For viability/cytotoxicity experiments we used 96 cell culture plates in which treatments were applied in triplicate (three times).
After 24 and 72 hours of incubation with the various AS-ODN treatment combinations the effect on cellular viability was studied. For that purpose we used the "LIVE/DEAD viability/cytotoxicity kit" from Molecular Probes®. This kit contains modified calcein (calcein-AM), a substance with silenced fluorescent calcein. When this substance penetrates viable cells, calcein is metabolized and recovers fluorescence, which is then measured in relative fluorescence units (RFU).
For the statistical analysis and graphic design we used the program "Analyzing Data with GraphPad Prism-1999", GraphPad Software Inc., San Diego CA©. After determining the fluorescence (RFU) emission with each dose (1, 5,10 and 20 µM) applied for 48 and 72 hours, a Student's t test was used to compare them with the RFU control group. A p < 0.05 was considered statistically significant. Although the statistical analysis was made using RFUs, our results are shown as rates of viability with respect to the control group, which presents a 100% viability rate, in order to facilitate reader comprehension (Table I).
RESULTS
Viability test with calcein-AM characterization
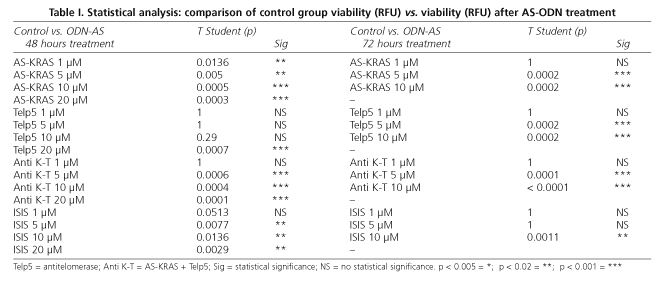
In order to know the optimal conditions for our study's technique, SW-480 cells were incubated with calcein for different periods of time, and the different fluorescent emissions were registered with a microplate cytofluorometer. Results (Fig. 1) show that a maximum of cellular fluorescence is reached after 40 minutes of incubation, so these were the conditions mantained for further studies.
Cell viability after 48 hours of treatment with AS-ODN
In figure 2 we show viability after 48 hours of treatment with AS-ODN depending on concentration:
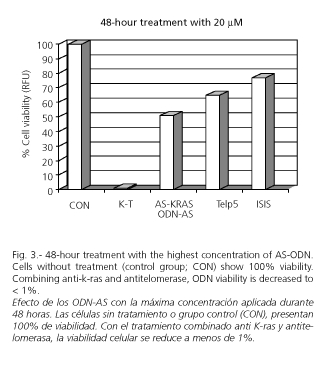
1. 20 µM concentration treatment:
-AS-KRAS decreases cell viability by 41%.
-Telp5 decreases cell viability by 26%.
-Combined AS-KRAS and Telp5 decrease cell viability by 99.6%.
-ISIS decreases cell viability by 14.52%.
2. 10 µM concentration treatment:
-AS-KRAS decreases cell viability by 32.3%
-Telp5 decreases cell viability by 0.6%.
-Combined AS-KRAS and Telp5 decrease cell viability by 34.65%.
-ISIS decreases cell viability by 6%.
3. 5 µM concentration treatment:
-AS-KRAS decreases cell viability by 32%.
-Telp5 decreases cell viability by 0%.
-Combined AS-KRAS and Telp5 decrease cell viability by 30%.
-ISIS decreases cell viability by 8%.
4. 1 µM concentration did not affect cell viability.
The effect of AS-ODN treatment at peak concentration for 48 hours is shown in figure 3.

Cell viability after 72 hours of treatment with AS-ODN
In figure 4 we show how cell viability decreases after 72 hours of treatment with AS-ODN:
1. 10 µM concentration treatment:
-AS-KRAS decreases cell viability by 56%.
-Telp5 decreases cell viability by 4%.
-Combined AS-KRAS and Telp5 decrease cell viability by 99.8%.
-ISIS decreases cell viability by 28.5%.
2. 5 µM concentration treatment:
-AS-KRAS decreases cell viability by 55.57%.
-Telp5 decreases cell viability by 54.68%.
-Combined AS-KRAS and Telp5 decrease cell viability by 63.07%.
-ISIS did not affect cell viability.
3. 1 µM concentrations did not affect cell viability.
Combined treatment with AS-KRAS and Telp5 caused a 100% mortality of tumoral cells.

DISCUSSION
In our study we tested the effect on cell viability of antisense therapy with oligodeoxiribonucleotides blocking the k-ras oncogene and telomerase gene in a human colorectal cancer line (SW480). Our results show that independent treatment with each AS-ODN can exert partially inhibiting effects (approximately 50%) on cell proliferation. Nevertheless, a combination of anti-k-ras and antitelomerase has a remarkable synergy that leads to a complete loss of tumoral cell viability (> 99.5%).
In vitro cytotoxicity tests are of great importance in the development of new biological agents with cytotoxic activity, especially if those procedures are of high sensitivity and avoid the use of conventional nuclide (usually Cr51) protocols. In our study we used a cytotoxicity test based on microplate cytofluorometry. It is based on fluorochrome calcein with chemically concealed fluorescence. After active cellular metabolization fluorescence is recovered, and this metabolization can only be done by viable cells.
Cytotoxicity studies with fluorometry show that all AS-ODNs tested achieved significant reductions in cell viability. Nevertheless, relevant differences in efficacy showed up. Of the two AS-ODNs tested against the k-ras oncogene, ISIS (designed to link mRNA's 5' terminal end of k-ras) showed a 30% inhibiting effect on cell viability. In equal conditions, AS-KRAS (designed to bind exon 1 of k-ras oncogene) had a higher inhibiting activity (55%). In both cases the maximum effect seems to have been reached, because after 72 hours of treatment period with 5 µM ODN concentrations viability does not decrease with higher concentrations. If we compare our results with those from other groups, a 20% viability reduction has been described by blocking k-ras with a cationic lipid leaded ODN after a 48-hour treatment period (17). Guan Chen's team achieved 70% of cell proliferation inhibition after 72 hours of treatment with anti k-ras ODN (14). In this same study ISIS 6957 was tested, and it only yielded 26% of inhibition in cellular growth with 400 nM after 72 hours of treatment. In their experiment they used a human lung fibroblast model and bladder cancer cells, so results are only partially comparable to ours, as we used another cell type for the tests.
Cell viability results with our antitelomerase ODN were similar in power and efficacy to those obtained with our anti-k-ras ODNs. Both 5 and 10 µM concentrations produced decreased viability in similar rates, which suggests that these doses are most specifically effective for Telp5.
Other studies (18-21) proved the possibility of blocking the telomerase gene with a phosphorotioate ODN. Herbert (22) demonstrated a 50% viability inhibition with 1 nanomolar lipid-vehiculized treatment, and also a viability reduction with non-lipid-vehiculized micromolar concentrations. In Herbert's experiment their antitelomerase ODN also binds the telomerase hTR subunit, but results are not comparable to ours for cultures were of breast cancer epithelial cells. Elayadi et al. (23) described telomerase gene blockade with a phosphorotioate ODN in a Jurkat T lymphocyte (leukemia) cell line. After 36 hours of treatment with 0.25 µM cellular activity decreased in 27%. A 2 µM dose decreased viability by 65%. ODNs were lipid-vehiculized with lipophectamine. Recently, Wong et al. (24) published their results using ODNs that link the telomerase hTR subunit in CRC, and they obtained varied results on viability reduction depending on the type of cell line tested (12-37%).
Our results in antisense gene therapy point out that AS-ODNs designed by our group are some of the most powerful ever described. Moreover, AS-KRAS acts earlier in time than Telp5. An explanation for this could be the different metabolical pathways followed by each AS-ODN. Nevertheless, what is of great relevance is the fact that our combined anti-k-ras and antitelomerase treatment has a synergic effect that leads to an inhibition of cell viability reaching > 99.5%. This opens an attractive alternative pathway in antisense therapy, specially considering that those results may be improved using transfection vectors. This, together with the improvement of cell transfection with AS-ODN (with a subsequent reduction in therapeutical dose) suggests the ability to simultaneously introduce combinations of AS-ODNs in cancer cells. Moreover, our results have been expressed as cell viability reduction rates. This is a good approximation to explain antisense therapy effects, but it would be desirable to quantify telomerase and k-ras expression, as well as treatment-derived proteins in further studies.
In summary, gene therapy with antisense oligonucleotides has been shown to be effective in our experience, especially the combination of anti k-ras and antitelomerase treatment, both implicated in CRC cell viability. In vivo applicability, usefulness and secondary effects need to be further established. Further studies will be needed to evaluate the in vivo efficacy of these AS-ODNs and their combinations using transfection vectors for the improvement of clinical applications.
ACKNOWLEDGEMENTS
This study was completely supported by an "Investigation Grant by the Spanish Gastroenterology Foundation". The authors want to thank this foundation for their support.
REFERENCES
1. Hermanek P, Wiebelt H, Staimmer D, Riedl S. Prognostic factors of rectum carcinoma. Experience of the German Multicentre Study SGCRC. German Study Group Colo-Rectal Carcinoma. Tumori 1995; 81 (Supl. 3): 60-4. [ Links ]
2. Hermanek P. Impact of surgeon's technique on outcome after treatment of rectal carcinoma. Dis Colon Rectum 1999; 42 (5): 559-62. [ Links ]
3. Umpleby HC, Fermor B, Symes MO, Williamson RC. Viability of exfoliated colorectal carcinoma cells. Br J Surg 1984; 71 (9): 659-63. [ Links ]
4. Solomon MJ, Egan M, Roberts RA, Philips J, Russell P. Incidence of free colorectal cancer cells on the peritoneal surface. Dis Colon Rectum 1997; 40 (11): 1294-8. [ Links ]
5. Ward RL, Todd AV, Santiago F, O'Connor T, Hawkins NJ. Activation of the K-ras oncogene in colorectal neoplasms is associated with decreased apoptosis. Cancer 1997; 79 (6): 1106-13. [ Links ]
6. Sakakura C, Hagiwara A, Tsujimoto H, Ozaki K, Sakakibara T, Oyama T, et al. Inhibition of colon cancer cell proliferation by antisense oligonucleotides targeting the messenger ARN of the Ki-ras gene. Anticancer Drugs 1995; 6 (4): 553-61. [ Links ]
7. Okada F, Rak JW, Croix BS, Lieubeau B, Kaya M, Roncari L, et al. Impact of oncogenes in tumor angiogenesis: mutant K-ras up-regulation of vascular endothelial growth factor/vascular permeability factor is necessary, but not sufficient for tumorigenicity of human colorectal carcinoma cells. Proc Natl Acad Sci USA 1998; 95 (7): 3609-14. [ Links ]
8. Dasí F, Marugán M, Lledó S, Cervantes A, García-Granero E, García-Conde J, et al. Telomerasa: biología e implicaciones clínicas. Hematol Citocinas Inmunoter Ter Cel 2002; 5 (2): 121-35. [ Links ]
9. Yoshida K, Sugino T, Goodison S, Warren BF, Nolan D, Wadsworth S, et al. Detection of telomerase activity in exfoliated cancer cells in colonic luminal washings and its related clinical implications. Br J Cancer 1997; 75 (4): 548-53. [ Links ]
10. Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL, et al. Specific association of human telomerase activity with immortal cells and cancer. Science 1994; 266 (5193): 2011-5. [ Links ]
11. Brown T, Aldous W, Lance R, Blaser J, Baker T, Williard W. The association between telomerase, p53, and clinical staging in colorectal cancer. Am J Surg 1998; 175 (5): 364-6. [ Links ]
12. Mercola D, Cohen JS. Antisense approaches to cancer gene therapy. Cancer Gene Ther 1995; 2 (1): 47-59. [ Links ]
13. Andreyev HJN, Ross PJ, Cunningham D, Clarke PA. Antisense treatment directed against mutated ki-ras in human colorectal carcinoma. Gut 2001; 48 (2): 230-7. [ Links ]
14. Chen G, Oh S, Monia BP, Stacey DW. Antisense oligonucleotides demonstrate a dominant role of c-ki-ras proteins in regulating the proliferation of diploid human fibroblasts. J Biol Chem 1996; 271 (45): 28259-65. [ Links ]
15. Pang JX, Chen XY, Xu W, Wu SG. Antisense Sp1 oligodeoxynucleotide decreases telomerase activity by inhibiting hTERT mRNA expression in Jurkat T cells. Acta Pharmacol Sin 2003; 24 (1): 91-6. [ Links ]
16. Pruzan R, Pongracz K, Gietzen K, Wallweber G, Gryaznov S. Allosteric inhibitors of telomerase: oligonucleotide N3'à P 5'phosphoramidates. Nucleic Acids Res 2002; 30 (2): 559-68. [ Links ]
17. Ross PJ, George M, Cunningham D, DiStefano F, Jervoise H, Andreyev H, et al. Inhibition of Kirsten-ras expression in human colorectal cancer using rationally selected kirsten-ras antisense oligonucleotides. Mol Cancer Ther 2001: 1 (1): 29-41. [ Links ]
18. Pang JX, Chen XY, Wu SG. Regulation of telomerase activity in Jurkat T cells by transcription factors Sp1 and Sp3. Di Yi Jun Yi Da Xue Xue Bao 2002; 22 (6): 481-5. [ Links ]
19. He DM, Zhang H. Telomerase gene antisense oligodoxynucleotide down-regulates telomerase activity in HL-60 cells. Ai Zheng 2002; 21 (10): 1070-4. [ Links ]
20. Wang X, Zhang Z, Xu Y, Chen S, Xiong W. Inhibition of telomerase activity and induction of apoptosis in lung cancer cell by human telomerase reverse transcriptase gene antisense oligodeoxynucleotide. Zhonghua Nei Ke Za Zhi 2002; 41 (3): 175-8. [ Links ]
21. Xie W, Lin M. Study on induction of leukemic cell apoptosis by antisense oligodeoxynucleotide of human telomerase RNA. Zhonghua Xue Ye Za Zhi 2001; 22 (5): 245-8. [ Links ]
22. Herbert BS, Pongracz K, Shay JW, Gryaznov SM, Shea-Herbert Brittney. Oligonucleotide N3'→P5'phosphoramidates as efficient telomerase inhibitors. Oncogene 2002; 21 (4): 638-42. [ Links ]
23. Elayadi AN, Demieville A, Wancewicz EV, Monia BP, Corey DR. Inhibition of telomerase by 2'-O- (2-methoxyethyl) RNA oligomers: effect of length, phosphorothioate substitution and time inside cells. Nucleic Acids Res 2001; 29 (8): 1683-9. [ Links ]
24. Wong SC, Yu H, Moochhala SM, So JB. Antisense telomerase induced cell growth inhibition, cell cycle arrest and telomerase activity down-regulation in gastric and colon cancer cells. Anticancer Res 2003; 23 (1 A): 465-9. [ Links ]

