










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkAnales del Sistema Sanitario de Navarra
versión impresa ISSN 1137-6627
Anales Sis San Navarra vol.31 no.3 Pamplona sep./dic. 2008
Nuevas estrategias terapéuticas en diabetes mellitus tipo 1
New therapeutic strategies for type 1 diabetes mellitus
M. Barajas1*, R. M. Príncipe2*, J. Escalada2, F. Prósper1, J. Salvador2
1. Departamento de Hematología y Área de Terapia Celular. Clínica Universitaria. Pamplona.
2. Departamento de Endocrinología y Nutrición. Clínica Universitaria. Pamplona.
* Ambos autores han contribuido por igual en el manuscrito final y, por tanto, comparten por igual la autoría del presente artículo.
Dirección para correspondencia
RESUMEN
El principal determinante del riesgo de complicaciones derivadas de la diabetes mellitus tipo 1 se debe a los altos niveles de glucosa en sangre mantenidos durante largo tiempo. Para conseguir un beneficio terapéutico en pacientes con diabetes mellitus es necesario desarrollar tratamientos que permitan de manera segura, efectiva y estable mantener la normoglucemia. Lamentablemente, el tratamiento de la diabetes mellitus tipo 1 mediante el aporte exógeno de insulina no es capaz de conseguir niveles estables de glucosa en sangre, de manera que con frecuencia se producen casos de severa hipoglucemia o hiperglucemia. Hasta la fecha la única solución para reestablecer de manera permanente la normoglucemia se consigue mediante el trasplante de páncreas o de islotes pancreáticos. Sin embargo, a medida que se incrementa el número de centros especializados en el trasplante de islotes, mayor es la necesidad de islotes para su trasplante. Así pues, el estudio de nuevas fuentes de células productoras de insulina así como de nuevos tratamientos que permitan preservar o incluso aumentar la masa de células beta en los pacientes con diabetes mellitus representa un objetivo de primera necesidad en este campo.
En este sentido, en la última década ha habido un avance significativo en el campo de la biología de las células madre. Sin embargo, la identificación de células apropiadas para la generación de nuevas células beta, además del desarrollo de técnicas para la caracterización de estas células, así como de ensayos y modelos animales apropiados para probar su capacidad de diferenciación tanto in vitro como in vivo son de vital importancia para la puesta en marcha de nuevas estrategias terapéuticas basadas en la aplicación de las células madre para el tratamiento de la diabetes mellitus tipo 1.
Palabras clave. Diabetes mellitus tipo 1. Células madre. Trasplante de páncreas. Islotes.
ABSTRACT
The main determinant of the risk of complications from type 1 diabetes mellitus is the total lifetime blood glucose levels. To impact on the health and quality of life of individuals with diabetes, safe and effective methods of achieving and maintaining normoglycemia are needed. Unfortunately, intensive insulin therapy does not achieve normal levels of blood glucose, is difficult to implement for many patients, and limited by the accompanying increased frequency of severe hypoglycemia. Hence, the only way at present to restore permanently normoglycemia without hypoglycemia is to provide the patient with additional beta-cells. This can be achieved by transplanting an intact pancreas, or by transplanting islets. The shortage of functional beta-cells from available donors is one of the major limiting factors for the treatment of diabetes by islet transplantation. Therefore, methods to preserve or even promote regeneration of the beta-cell mass are dearly needed.
Significant progress has been made over the last decade in stem cell biology. However, the quest for identification of stem cells has been hampered by the lack of appropriate research tools including assays that allow assess their differentiation potential in vitro and in vivo. Therefore, new techniques are necessary in order to develop new therapeutic strategies based on stem cells for the treatment of diabetes mellitus type 1.
Key words. Type 1 Diabetes mellitus. Stem cells. Pancreas transplantion. Islets.
Introducción
La incidencia de la diabetes mellitus tipo 1 (DM1) muestra una marcada diferencia geográfica y un incremento anual en casi todos los países, teniendo una mayor incidencia en los países escandinavos y Cerdeña. La susceptibilidad a desarrollar DM1 parece estar asociada a factores genéticos múltiples, aunque sólo el 25-50% de los pacientes presentan antecedentes familiares1.
La DM1 tiene lugar como consecuencia de una destrucción autoinmune de las células beta presentes en los islotes de Langerhans del páncreas. Estas células son las encargadas de la producción y secreción de insulina en respuesta al estímulo de la glucosa. La supervivencia de los pacientes con DM1 depende de la inyección diaria de insulina en varias dosis. Los resultados obtenidos con el trasplante de islotes han demostrado que esta estrategia es eficaz para el tratamiento de la DM1. Pacientes sometidos a esta intervención han conseguido vivir durante más de dos años sin tener que tratarse con insulina diariamente2. Sin embargo, esta aproximación terapéutica presenta una serie de inconvenientes entre los que cabe destacar aquellos que derivan de la medicación inmunosupresora, así como la necesidad de trasplantar islotes provenientes de 2 ó incluso 3 donantes para alcanzar la normoglucemia. Por tanto, es necesario mejorar el proceso de aislamiento de los islotes, su rendimiento, así como la búsqueda de nuevos medicamentos inmunosupresores u otras estrategias que permitan conseguir la tolerancia del trasplante. Por otra parte, teniendo en cuenta el bajo número de donantes de islotes respecto a la incidencia de la DM1, es necesario explorar vías alternativas para hacer posible la aplicación de esta terapia. Entre estas alternativas la más prometedora se basa en la utilización de las células madre, tanto para generar nuevas células beta que puedan ser trasplantadas en pacientes con DM1 de igual manera que se ha hecho con el trasplante de islotes, como para bloquear la respuesta autoinmune responsable de la generación de la enfermedad en el caso de la DM1.
Trasplante de páncreas
El trasplante de páncreas (TP) es la única terapia capaz de conseguir un control metabólico estricto, con valores fisiológicos de hemoglobina glicosilada. Actualmente, gracias a la mejoría de la técnica quirúrgica y al desarrollo de nuevos regímenes de inmunosupresión, el TP ha alcanzado un alto porcentaje de éxitos3,4. Las variantes del TP experimentadas hasta la actualidad son tres: el trasplante simultáneo riñón-páncreas (TRP), que supone casi el 90% del total; el trasplante de páncreas después de riñón (TPDR), que representa en torno el 10%, y el trasplante de páncreas aislado (TPA), muy poco utilizado.
Receptor
La selección de los candidatos debe ser muy rigurosa, ya que el éxito del TP se debe en gran medida a la selección de los pacientes.
Indicaciones
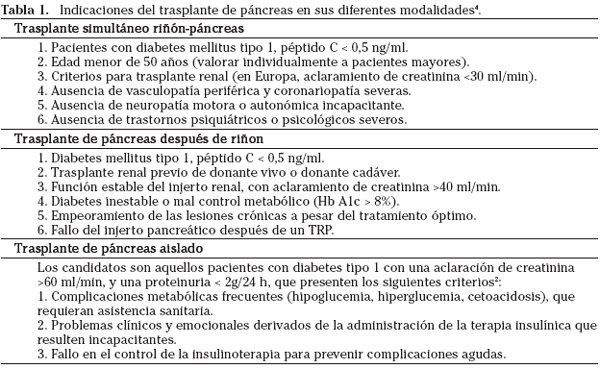
Las indicaciones para las diferentes modalidades de TP se describen en la tabla 1.
Contraindicaciones
Se consideran contraindicaciones absolutas: infección activa, neoplasia, alteraciones de la coagulación, coronariografía con lesiones no corregibles, fracción de eyección < 50%, infarto de miocardio reciente, obesidad severa, incumplimiento de tratamiento previo, drogadicción o alcoholismo activos y enfermedad psiquiátrica.
Son contraindicaciones relativas: IMC > 30 kg/m2, hemorragia retiniana reciente, fumador activo, serología positiva VHC y VHB.
Donante de páncreas
El donante ideal es un individuo fallecido por traumatismo craneoencefálico o por hemorragia intracraneal secundaria a malformaciones vasculares. El estado de los vasos ilíacos es fundamental para una correcta reconstrucción vascular y su correcto funcionamiento posterior.
El rango de edad recomendado oscila entre 10 y 45 años. La edad superior a 45 años no es una contraindicación, pero sí merece una valoración rigurosa.
El donante debe tener un peso mínimo de 30 kg, ya que el páncreas es un órgano de bajo flujo, y en aquellos páncreas con vasos de menor calibre se favorece la trombosis vascular y por tanto, la pérdida del injerto. Un IMC > 30 kg/m2 es una contraindicación ya que suele implicar una degeneración grasa del páncreas, con un aumento en la incidencia de pancreatitis, trombosis e infección. Con la introducción de la solución de Wisconsin, el tiempo de isquemia fría aceptado para que el injerto de páncreas pueda funcionar con normalidad es hasta de 30 horas. En los últimos años el tiempo medio de isquemia fría ha sido de 15 horas para el trasplante aislado de páncreas y de 13 horas para el simultáneo páncreas-riñón5.
La muerte por intoxicación no constituye una contraindicación salvo en el caso del metanol. La hiperglucemia iatrogénica tampoco, en ausencia de historia de insuficiencia endocrina. La elevación aislada de los niveles de amilasa plasmática, en ausencia de otras comorbilidades no constituye una contraindicación para la donación.
Complicaciones quirúrgicas
Las complicaciones específicas derivadas de esta cirugía son las siguientes: vasculares, intraabdominales, pancreatitis del injerto, fuga anastomótica y las relacionadas con el drenaje vesical.
Complicaciones vasculares
Trombosis. Es la más frecuente: 6% en TRP, 7% en TPDR, y un 9% en TPA. Supone casi la mitad de los fallos no inmunológicos del injerto y se manifiesta en las dos primeras semanas hasta en el 90% de los casos. El paciente suele presentar dolor abdominal y el diagnóstico se confirma mediante un eco-doppler. La pancreatectomía del injerto suele ser el tratamiento habitual.
Hemorragia intraabdominal. Un factor de riesgo importante es la utilización de anticoagulación.
Pseudoaneurisma micótico. Se produce siempre en el contexto de una infección intraabdominal.
Infecciones intraabdominales
Pueden reducir significativamente la supervivencia del injerto y del paciente. La mayoría de los centros utilizan el fluconazol como profilaxis antifúngica y la combinación del imipenem o la piperacilina junto con la vancomicina como profilaxis bacteriana.
Pancreatitis del injerto
Se manifiesta mediante una hiperamilasemia prolongada. El octreótide puede ser utilizado para la profilaxis o el tratamiento de esta complicación.
Fuga anastomótica
Se diagnostica por TC abdominal con inyección de contraste vesical en el caso del trasplante con drenaje vesical, y por TC abdominal con contraste oral en el caso del trasplante con drenaje entérico. Es necesaria la reparación primaria de la fuga anastomótica hasta en un 66% de los casos, o en su defecto la conversión entérica. En caso de fuga por drenaje entérico la laparotomía exploratoria es casi siempre necesaria para un adecuado manejo del paciente.
Complicaciones relacionadas con el drenaje vesical
Las complicaciones más serias son las urológicas, en orden de frecuencia: infección recurrente del tracto urinario, hematuria crónica, fuga urinaria y lesiones uretrales. La mayoría de los pacientes con complicaciones uretrales suelen precisar la conversión entérica para la desaparición de los síntomas.
Resultados
Según el Registro Internacional de Trasplante de Páncreas, en el TRP, la supervivencia al año del paciente, el injerto pancreático y renal ha mejorado de un 90, 74 y 83% respectivamente en el período 1987-1989, a un 95, 83 y 88% durante el período 1996-2000, en los pacientes tratados en EEUU6. Mientras que en el TPDR y en el TPA la supervivencia del injerto de páncreas al año ha sido en este último período de 73 y 70% respectivamente7.
Supervivencia del paciente
Diferentes estudios han demostrado que existe una mayor supervivencia en pacientes tratados mediante TRP frente a aquellos tratados mediante trasplante aislado de riñón.
La ausencia de ensayos clínicos controlados representa una barrera para comprender y comparar los riesgos y beneficios del TPA y el TPDR. Sin embargo, hay datos sobre la mortalidad relacionada con el postoperatorio, sin aparentes beneficios de supervivencia a largo plazo8.
Supervivencia del injerto renal
La supervivencia del injerto renal en el TRP en pacientes con DM1 es comparable a la supervivencia del injerto renal en trasplante aislado de riñón de cadáver. Sin embargo, diversas hipótesis sostienen que existe un claro beneficio para el riñón transplantado si existe una buena función del injerto pancreático. Los pacientes con TRP observados a los 2 años y medio tras la cirugía, comparados con aquellos pacientes con trasplante aislado de riñón no presentan engrosamiento de la membrana basal glomerular (92% vs 35%) ni expansión del volumen mesangial (82% vs 12%)9. Además, se ha observado que la progresión de la glomerulopatía en pacientes trasplantados de riñón puede detenerse a los dos años tras un trasplante de páncreas diferido.
Efectos metabólicos
El TP permite restaurar el nivel de glucosa en sangre y la respuesta bifásica de la insulina tras la estimulación con glucosa, obteniendo una normalización de la hemoglobina glicosilada en semanas o meses tras la cirugía.
Aunque pueden ocurrir episodios de hipoglucemia por hiperinsulinismo, el coma hipoglucémico es excepcional. La aparición de hiperinsulinismo se ha asociado al drenaje venoso sistémico, a una disminución del aclaramiento de insulina por el hígado, a los fármacos anticalcineurínicos y los esteroides, y a una supresión incompleta de insulina debida a la denervación existente en el injerto pancreático.
Retinopatía
En la mayoría de los estudios, los pacientes sometidos a un TP presentan retinopatía proliferativa en el momento del trasplante. Por ello, se tratan mediante coagulación panretiniana con láser. Esto podría justificar la estabilización de la retinopatía diabética tras el trasplante6-10.
Neuropatía
Los pacientes diabéticos con neuropatía moderada que se someten a un TP tienen una mayor supervivencia que aquellos no sometidos a trasplante o con un fallo precoz del mismo. Está descrita la recuperación de la conducción nerviosa sensitivomotora, con desaparición de la polineuropatía dolorosa y el síndrome de piernas inquietas en un corto período de tiempo10,11.
Enfermedad cardiovascular
Se ha observado una progresión de la enfermedad establecida macrovascular a pesar de la realización satisfactoria del TP. Su influencia sobre la evolución de la enfermedad vascular asintomática está aún por demostrar. Sin embargo, el TP puede tener un efecto positivo en la función cardíaca y se consigue una reducción en la presión arterial, lo que beneficiaría a los pacientes con nefropatía diabética establecida.
Calidad de vida
En pacientes diabéticos insulinotratados, la mejora en la calidad de vida y la prevención o estabilización de la morbilidad y mortalidad asociada a las complicaciones de la diabetes hacen del TP una importante opción terapéutica12.
Trasplante de islotes
El trasplante de islotes, aunque desarrollado en 1967, ha permanecido casi en el olvido hasta que en el año 2000, Shapiro y col en la Universidad de Alberta, en Edmonton, describieron un nuevo protocolo para el trasplante de islotes con el que consiguieron la insulinoindependencia al año del trasplante en hasta el 80% de pacientes13.
Estos resultados han sido parcialmente reproducidos en un estudio internacional, el International Network Trial (INT), en el que el 58% de los pacientes consiguieron la independencia de la insulina, aunque con amplias variaciones entre centros (desde el 100% al 0% de pacientes) que obedecían fundamentalmente a la experiencia previa del grupo en trasplante de islotes13. Sin embargo, los datos de seguimiento a más largo plazo muestran un deterioro de la función del injerto, de forma que a los 2 años del trasplante tan sólo el 24% de pacientes mantenían la insulinoindependencia en el estudio INT, porcentaje que el grupo de Edmonton ha descrito que se reduce hasta el 10% a los 5 años14,15. Aunque estos resultados han supuesto una cierta decepción, es importante señalar que, en cerca del 70% de los pacientes, el injerto muestra una función beta residual, lo que permite a estos pacientes mantener un control glucémico óptimo con dosis muy bajas de insulina, con gran estabilidad metabólica (indicado por una reducción en la amplitud de las excursiones glucémicas), y sin episodios de hipoglucemias graves.
Los cambios más importantes introducidos por el protocolo de Edmonton han sido13, 1:
El uso de una fuerte inmunosupresión en la que se prescindió de los glucocorticoides y se incluyó sirolimus, bajas dosis de tacrolimus y un anticuerpo monoclonal frente al receptor de la interleuquina-2 (daclizumab).
La eliminación durante el proceso de aislamiento de los islotes del uso de productos que pudieran contener xenoproteínas con la intención de reducir la destrucción inmediata tras el trasplante.
La minimización del tiempo de isquemia, trasplantando los islotes inmediatamente después de su aislamiento, aunque este aspecto se ha abandonado en estudios posteriores siendo común en la actualidad trasplantar los islotes tras 2-3 días de cultivo.
El trasplante de una masa beta superior a la usada previamente, con una media de más de 4.000 islotes-equivalente por kilogramo de peso y la repetición del trasplante en 2-3 ocasiones en caso de detectar glucemias superiores a 200 mg/dl en las semanas posteriores al trasplante.
Trasplantar a una población receptora distinta de las usadas hasta entonces, que no presentaba insuficiencia renal, en su mayoría pacientes con episodios repetidos de hipoglucemia grave.
Aunque el proceso no ha tenido mortalidad y su morbilidad es baja comparada con el trasplante de órganos, el procedimiento no está exento de efectos adversos, algunos de ellos graves. En relación al procedimiento quirúrgico son destacables la presencia de sangrado grave en un 10% de pacientes, que en algún caso ha requerido nueva cirugía, la trombosis de la porta o alguna de sus ramas (6%), y la elevación transitoria de enzimas hepáticos. Como eventos adversos no graves más frecuentes se describen las úlceras bucales (92%), anemia (81%), leucopenia (75%), diarrea (64%), cefalea (56%), neutropenia (53%), naúseas (50%), vómitos (42%), acné (39%) y astenia (39%). Hasta el momento no se ha descrito la aparición de neoplasias malignas en los pacientes con trasplante de islotes y tratamiento inmunosupresor.
Así pues, a día de hoy, el trasplante de islotes está limitado a un escaso número de pacientes y centros. Para poder generalizar esta opción a la mayoría o al menos a un número significativo de pacientes, la investigación debe resolver problemas fundamentales como son la mejora de las condiciones técnicas del aislamiento de islotes para aumentar el rendimiento y la reproducibilidad del proceso, así como la funcionalidad de los islotes obtenidos15.
Finalmente, para poder hacer frente a la enorme demanda debida al potencial número de receptores del trasplante es preciso disponer de sistemas de expansión de los islotes in vitro, o de fuentes alternativas de células productoras de insulina, como por ejemplo la generación de células beta a partir de células madre adultas.
Tratamiento de la diabetes con células madre
Hace tan sólo una década desde la aparición de la terapia celular con células madre como una nueva área de investigación. Hasta la fecha se consideraba que las células poseían una capacidad de división y diferenciación limitada. Sin embargo, este dogma está cambiando gracias a recientes resultados publicados acerca de la pluripotencialidad de las células madre. Así pues, células procedentes de un determinado origen han sido capaces de dar lugar a células con características propias de otro linaje. La posibilidad de trasplantar estas células en órganos, con el objeto de sustituir un tejido dañado, ha impulsado notablemente el estudio del fenómeno conocido como plasticidad celular y su potencial aplicación en situaciones patológicas. El beneficio terapéutico observado después del trasplante de células madre ha sido probado con éxito tanto a nivel experimental como, en algunos casos, clínicamente. Sin embargo, el grado de contribución de estas células al tejido trasplantado ha resultado ser menor de lo esperado, de manera que cada vez hay más evidencias que sugieren la existencia de mecanismos paracrinos que jugarían un papel muy importante en el beneficio terapéutico observado. El estudio en profundidad de estos efectos tróficos inducidos por las células madre trasplantadas podría ser de gran relevancia para el tratamiento de tejidos dañados, potenciando su regeneración a partir de las células endógenas no dañadas presentes en el órgano afectado. Por otra parte, el desarrollo de métodos más sofisticados de trasplante de células combinado con técnicas de bio-ingeniería podría mejorar notablemente el potencial de las células madre.
Por otra parte, la puesta en escena de un concepto ampliamente estudiado, la reprogramación celular, está cambiando notablemente el panorama actual en el campo de la investigación con células madre. Recientes estudios demuestran que es posible obtener células con capacidad de expansión y diferenciación similar al observado en las células madre embrionarias, mediante la transferencia de cuatro genes, tales como Oct3/4, Sox2, Klf4, y c-Myc (siendo la utilización de este último opcional), partiendo de células adultas, tales como por ejemplo fibroblastos derivados de la dermis.
A continuación mostramos algunos de los tipos de células madre que actualmente se están estudiando a nivel experimental para el tratamiento de la diabetes.
Tipos de células madre
Células madre embrionarias
Las células madre embrionarias (ESCs, del inglés embryonic stem cells) derivan de la masa interna del blastocisto. Una vez aisladas se expanden mediante su cultivo sobre fibroblastos murinos embrionarios cuya función es servir como soporte para su crecimiento. Las ESCs son capaces de formar cuerpos embrioides, así como diferenciarse hacia células propias de las tres capas embrionarias: ectodermo, endodermo y mesodermo. Numerosos grupos de investigación han demostrado que las ESCs son capaces de diferenciarse hacia células productoras de insulina in vitro16-18. Incluso diferentes autores han conseguido enriquecer el número de células productoras de insulina a partir de ESCs que se diferencian espontáneamente, utilizando técnicas como:
La selección mediante manipulación del medio de cultivo18,19.
La sobreexpresión de factores de transcripción claves tales como Pdx-1 o Pax420,21.
La selección mediante la utilización de resistencia a antibióticos dirigidos por los promotores para Nkx6.1 o insulina22,16.
La diferenciación hacia células productoras de insulina utilizando factores de crecimiento solubles, matrices celulares y el contacto célula-célula23.
Células madre derivadas de la médula ósea
Los resultados recientemente publicados sobre el potencial de diferenciación de las células madre derivadas de la médula ósea están cambiando la visión clásica de las células madre específicas de tejido, de manera que se sabe que algunas poblaciones celulares presentes en la médula ósea son capaces de abandonar su patrón de diferenciación celular característico y diferenciarse hacia tejidos típicamente no-hematopoyéticos24. Este fenómeno está favorecido, en cierta medida, por la presión selectiva inducida por el tejido dañado. Estudios recientes demuestran que es posible detectar la expresión de insulina en tejidos en los que habitualmente ésta no tiene lugar, tales como la médula ósea, el tejido adiposo o el músculo esquelético25,26. Estos datos sugieren que la hiperglicemia es capaz de inducir la expresión de insulina en poblaciones de células presentes en la médula ósea. Incluso se han llegado a detectar células cuyo origen es la médula ósea que expresan insulina en el tejido acinar, aunque nunca de manera específica en los islotes pancreáticos26.
Otros autores afirman que algunas células presentes en la médula ósea son capaces de transdiferenciarse dando lugar a células beta funcionales. Ianus y col27 han demostrado, utilizando el sistema Cre-loxP, que las células derivadas de la médula ósea contribuyen aproximadamente entre 1,7 y 3% a la formación de nuevas células beta. Además, estos autores demuestran que estos resultados no se deben a procesos de fusión celular, indicando que las células adultas presentes en la médula ósea son capaces, aunque con baja frecuencia, de diferenciarse directamente hacia células beta en los islotes pancreáticos.
Células madre mesenquimales derivadas de la médula ósea
Las células madre mesenquimales (MSCs, del inglés Mesenchymal Stem Cells) de la médula ósea se han definido clásicamente por su capacidad para dar lugar a tejidos tales como el cartílago o hueso28. Estas células han despertado un gran interés debido a su prometedor potencial terapéutico, porque además de su capacidad de diferenciarse hacia los citados tejidos, algunos autores han demostrado que además son capaces de adoptar fenotipos propios de tejidos ectodérmicos y endodérmicos in vitro e in vivo28,29. Las MSCs derivadas de médula ósea murina han sido recientemente diferenciadas hacia células productoras de insulina in vitro e incluso han demostrado ser capaces de revertir la hiperglicemia en un modelo murino de diabetes en ratón30. Además, se ha demostrado que es posible diferenciar MSCs derivadas de la médula ósea humana hacia células productoras de insulina cuando se cultivan en el microambiente adecuado y utilizando mecanismos que implican factores de transcripción importantes para el desarrollo de las células beta, tales como Pdx131.
Pese al escaso conocimiento sobre el papel de las MSCs en el mantenimiento de la homeostasis o su implicación en el desarrollo de determinadas patologías, durante los últimos años se ha investigado de manera extensiva sobre el papel antiproliferativo, inmunomodulador y antiinflamatorio de estas células. Las células MSCs son capaces de bloquear la proliferación de linfocitos alogénicos in vitro, incluso después de haber sido tratadas con IFN-gamma para inducir la expresión de moléculas de Clase II o transfectadas con moléculas coestimuladoras como B7-1 o B7-2. De manera similar, se sabe que las MSCs no son lisadas por linfocitos T citotóxicos o células NK (del inglés Natural Killer). Esta capacidad de supresión parece estar mediada, al menos en parte, por factores solubles secretados por las MSCs, entre los que se encuentra el TGF-beta32, el HGF o la prostagladina E2.
In vivo, las MSCs son capaces de prolongar el tiempo de rechazo de trasplantes de piel, así como mejorar la funcionalidad renal, neural y pulmonar en diferentes modelos experimentales animales. Estos efectos se cree que son debidos a una acción paracrina derivada de las MSCs que lleva a modificar el entorno en el sitio de la lesión desde una situación que en condiciones normales debería ser proinflamatoria perjudicial hacia una situación antiinflamatoria beneficiosa.
En humanos, más de 100 pacientes han sido transplantados con MSCs33, en muchos de los casos, derivadas de donantes emparentados y utilizadas con la finalidad de mejorar el trasplante de células madre hematopoyéticas para el tratamiento de leucemias. La infusión de células MSCs ha sido bien tolerada en todos los casos, sin la aparición de ningún efecto adverso. En un paciente de 9 años de edad se observó una protección frente a una reacción inmunológica derivada del trasplante de células madre hematopoyéticas alogénicas para el tratamiento de una leucemia34. Las MSCs también han sido ensayadas con éxito para el tratamiento de enfermedades autoinmunes como la enfermedad injerto-contra-huésped (GvHD, del inglés Graft versus Host Disease), donde la infusión de MSCs derivadas de donantes emparentados ha demostrado ser eficaz revertiendo con éxito el daño provocado por esta enfermedad en el hígado e intestino de estos pacientes 34,35.
Recientemente nuestro grupo de investigación está llevando a cabo experimentos cuya finalidad última es determinar una posible aplicación de las MSCs para el tratamiento de la diabetes autoinmune. Nuestro principal objetivo consiste en estudiar si las MSCs pueden ejercer un efecto protector y así frenar la aparición de diabetes en el modelo de diabetes autoinmune tipo 1 que desarrolla el ratón NOD. Para ello trasplantamos MSCs derivadas de 2 cepas de ratones diferentes con el objeto de comparar el trasplante autólogo (células MSCs derivadas de ratón NOD) respecto al trasplante alogénico (células MSCs derivadas de ratón C57Bl/6). Nuestros resultados preliminares indican que el tratamiento de ratones NOD con MSCs protege del desarrollo de diabetes, retrasando en muchos casos la aparición de la enfermedad.
Células derivadas del bazo
El grupo liderado por el Dr. Faustman demostró en el año 200336 que era posible reestablecer la normoglicemia en el modelo de diabetes autoinmune que se desarrolla en ratones NOD mediante el trasplante de células derivadas del bazo (esplenocitos) y previo tratamiento con adyuvante de Freund completo (CFA, del inglés Complete Freunds Adjuvant). Estos investigadores pusieron de manifiesto la presencia de células procedentes de los animales donantes en los islotes y ductos pancreáticos. Además, estos autores también determinaron que los islotes recién formados se habían originado mediante diferenciación a partir de los esplenocitos infundidos y no mediante un proceso de fusión celular entre las células trasplantadas y las células beta residuales endógenas. Sin embargo, estudios recientes utilizando la misma aproximación no han sido capaces de reproducir tales resultados37-39, mostrando sin embargo que el tratamiento con CFA es suficiente para reinstaurar los niveles normales de glicemia mediante un proceso de regeneración de las células beta endógenas, de manera, que no queda claro el papel que podrían desempeñar los esplenocitos en la regeneración de los islotes.
Células madre del hígado con potencial pancreático
Existen en la literatura varios artículos que sugieren que el hígado, por derivar de la misma capa embrionaria que el páncreas (el endodermo), podría ser un candidato ideal para ser utilizado para el tratamiento de la diabetes mediante su transdiferenciación hacia células beta. Así por ejemplo, se sabe que la expresión de genes importantes para el desarrollo del páncreas (como por ejemplo Pdx-1) en hepatocitos, es capaz de inducir la expresión de marcadores maduros de célula beta, incluyendo la expresión de insulina40. De manera similar, la sobreexpresión de betacelulina y NeuroD/Beta2 ha demostrado ser una aproximación eficaz para convertir a un hepatocito en un célula productora de insulina, capaz incluso de revertir la diabetes inducida en ratones de experimentación por estreptozotocina (STZ)41. Por otra parte, las células ovales aisladas del hígado de ratas sometidas a un daño son capaces de diferenciarse formando estructuras que simulan islotes pancreáticos capaces incluso de secretar insulina en respuesta a concentraciones altas de glucosa42.
Células madre del páncreas
Banerjee and Bhonde43 han podido aislar células madre a partir de células beta presentes en los islotes pancreáticos de ratones tratados con STZ utilizando la combinación del factor de crecimiento derivado de queratinocitos (KGF, del inglés Keratinocyte Growth Factor) junto con el factor derivado de fibroblastos (FGF, del inglés Fibroblast Growth Factor). Los nuevos islotes formados siguiendo este procedimiento fueron capaces de expresar Pdx-1, así como secretar insulina en respuesta a glucosa. Estos resultados sugieren que los islotes a los que se les elimina de manera selectiva las células beta mantienen células con capacidad para generar el islote completo, incluyendo las células beta perdidas43.
Evidencias indirectas, examinando la distribución de células que expresan los marcadores Pdx-1 y Ngn3 tras la administración de STZ, sugieren la presencia de progenitores de célula beta en el interior de los islotes pancreáticos44.
Suzuki y col45 han sido capaces de aislar células que expresan el marcador c-Met a partir de páncreas de ratón neonato y adulto46, así como inducir la aparición de estructuras con apariencia similar a los islotes a partir de células epiteliales ductales del páncreas que expresan c-Met47.
Monocitos
Ruhnke y col han demostrado que es posible obtener células productoras de insulina a partir de monocitos CD14+ derivados de sangre periférica mediante un protocolo de diferenciación consistente en una combinación del factor estimulador de colonias de macrófagos (M-CSF, del inglés Macrophage Colony-Stimulating Factor) e Interleucina 348. La acción sinérgica de ambas citoquinas, sumado a la plasticidad fenotípica de los monocitos, es la responsable de la diferenciación de estas células de origen mielomonocítico hacia células beta. Los autores demuestran que es posible utilizar estas células para tratar ratones diabéticos, siendo capaces de normalizar los niveles de glucosa en sangre cuando son trasplantados bajo la cápsula renal de ratones diabéticos inducidos por STZ48.
Células madre pluripotentes inducidas (iPS).
Recientemente se ha descrito un nuevo procedimiento para obtener células madre pluripotentes, es decir, células con capacidad de diferenciación hacia tejidos de las tres capas embrionarias, partiendo de células madre adultas, tales como fibroblastos derivados de la piel49,50. Estas células reciben el nombre de células madre pluripotentes inducidas (iPS, del inglés Induced Pluripotent Stem cells).
Las iPS suponen un avance muy significativo en el campo de las células madre ya que hasta la fecha la capacidad de diferenciación que se les atribuía a las células adultas era muy limitada. La aplicación de novedosa tecnología, que consiste en la transferencia de cuatro genes (Oct3/4, Sox2, Klf-4 y c-Myc), abre un nuevo campo de investigación: la reprogramación de células adultas. Una mayor compresión de los fenómenos que subyacen al proceso de reprogramación, así como una mejora en los protocolos de generación de iPS, hace pensar que en un futuro no muy lejano será posible el tratamiento de enfermedades para las cuales hasta la fecha no hay curación alguna51. Respecto a la enfermedad que nos ocupa, no sólo se ha conseguido generar iPS a partir de tejido pancreático52 sino también diferenciar con éxito iPS hacia células productoras de insulina 53,54.
Conclusiones
Aunque la diabetes puede ser tratada mediante el aporte exógeno de insulina, el trasplante de islotes pancreáticos proporciona un mejor control glucémico, evitando la administración diaria de insulina. Sin embargo, limitaciones en la cantidad de tejido accesible para su trasplante, ha impedido la aplicación de esta estrategia terapéutica para el tratamiento de la DM1. En este sentido, las células madre constituyen una alternativa muy prometedora, ya que podrían suplir las células beta necesarias para el tratamiento de la enfermedad sin la necesidad de someter a los pacientes a severos regímenes de inmunosupresión. El objetivo final debería ser la creación, no sólo de células beta, sino de islotes funcionales donde estén presentes el resto de tipos celulares. Encontrar la población de células madre adecuada para este propósito es un objetivo de primera necesidad.
Por otra parte, la aplicación de propiedades hasta la fecha desconocidas atribuidas a determinados tipos de células madre, tales como la inmunosupresión inducida por las MSCs, podría ser de gran utilidad para su combinación junto con células beta o islotes derivados a partir de células madre. La combinación de ambas estrategias previsiblemente mejoraría los resultados que se obtengan con el trasplante de células madre derivadas hacia células beta, permitiendo incluso disminuir el número de islotes o células beta necesarias para el tratamiento de la diabetes. De hecho, el desarrollo de nuevos regímenes inmunosupresores en el Protocolo de Edmonton ha supuesto una notable mejora en la aplicación de esta estrategia terapéutica.
Por último, la entrada en escena de un nuevo tipo celular, las iPS, promete revolucionar el campo de las células madre en el tratamiento de la DM1. Sin embargo, hará falta esperar al menos una década hasta que podamos ver los primeros resultados tras la implantación de células beta o islotes derivados de iPS en pacientes diabéticos.
Bibliografía
1. Adeghate E, Schattner P, Dunn E. An update on the etiology and epidemiology of diabetes mellitus. Ann N Y Acad Sci 2006; 1084: 1-29. [ Links ]
2. Shapiro AMJ, Ricordi C, Hering BJ, Auchincloss H, Lindblad R, Robertson RP et al. International trial of the edmonton protocol for islet transplantation. N Engl J Med 2006; 355: 1318-1330. [ Links ]
3. Ming CS, Chen ZH. Progress in pancreas transplantation and combined pancreas-kidney transplantation. Hepatobiliary Pancreat Dis Int 2007; 6: 17-23. [ Links ]
4. Montiel MC, Pardo F, Rotellar F, Valentí V, Pastor C, Álvarez Cienfuegos J. Trasplante pancreático. An Sist Sanit Navar 2006; 29 (Supl 2): 113-124. [ Links ]
5. Sutherland DER, Gruessner RWG, Dunn DL, Matas AJ, Kandaswamy R, Mauer SM et al. Lessons learned from more than 1000 pancreas transplants at a single institution. Ann Surg 2001; 233: 463-501. [ Links ]
6. Kahl A, Bechstein WO, Frei U. Trends and perspectives in pancreas and simultaneous pancreas and kidney transplantation. Curr Opin Urol 2001; 11: 165-174. [ Links ]
7. International Pancreas Transplant Registry. Ed. B. J. Bland, Minneapolis, Minesota, USA. http://www.iptr.umn.edu. [ Links ]
8. Nathan DM. Isolated pancreas transplantation for type 1 diabetes. JAMA 2003; 290: 2861-2863. [ Links ]
9. Fioretto P, Steffes MW, Sutherland DER, Goetz FC, Mauer M. Reversal of lesions of diabetic nephropathy after pancreas transplantation. N Engl J Med 1998; 339: 69-71. [ Links ]
10. Ryan EA, Bigam D, Shapiro J. Current indications for pancreas or islet transplant. Diabetes, Obes Metab 2006; 8: 1-7. [ Links ]
11. Navarro X, Sutherland DE, Kennedy WR. Long-term effects of pancreatic transplantation on diabetic neuropathy. Ann Neurol 1997; 42: 727-736. [ Links ]
12. Demartines N, Schiesser M, Clavien PA. An evidence-Based Analysis of simultaneous pancreas-kidney and pancreas transplantation alone. Am J Transplant 2005; 5: 2688-2697. [ Links ]
13. Shapiro AMJ, Lakey JRT, Ryan EA, Korbutt GS, Toth E, Warnock GL et al. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N Engl J Med 2000; 343: 230-238. [ Links ]
14. Ryan EA, Paty BW, Senior PA, Bigam D, Alfadhli E, Kneteman NM et al. Five-year follow-up after clinical islet transplantation. Diabetes 2005; 54: 2060-2069. [ Links ]
15. Robertson RP. Islet transplantation as a treatment for diabetes. A work in progress. N Engl J Med 2004; 350: 694-705. [ Links ]
16. Soria B, Roche E, Berná G, león-quinto T, Reig JA, Martín F. Insulin-secreting cells derived from embryonic stem cells normalize glycemia in streptozotocin-induced diabetic mice. Diabetes 2000; 49: 157-162. [ Links ]
17. Assady S, Maor G, Amit M, Itskovitz-Eldor J, Skorecki KL, TzuKerman M. Insulin production by human embryonic stem cells. Diabetes 2001; 50: 1691-1697. [ Links ]
18. Lumelsky N, Blondel O, Laeng P, Velasco I, Ravin R, McKay R. Differentiation of embryonic stem cells to insulin-secreting structures similar to pancreatic islets. Science 2001; 292: 1389-1394. [ Links ]
19. Hori Y, Rulifson IC, Tsai BC, Heit JJ, Cahoy JD, Kim SK. Growth inhibitors promote differentiation of insulin-producing tissue from embryonic stem cells. Proc Natl Acad Sci USA 2002; 99: 16105-16110. [ Links ]
20. Miyazaki S, Yamato E, Miyazaki J. Regulated expression of pdx-1 promotes in vitro differentiation of insulin-producing cells from embryonic stem cells. Diabetes 2004; 53: 1030-1037. [ Links ]
21. Blyszczuk P, Czyz J, Kania G, Wagner M, Roll U, St-Onge L et al. Expression of Pax4 in embryonic stem cells promotes differentiation of nestin-positive progenitor and insulin-producing cells. Proc Natl Acad Sci USA 2003; 100: 998-1003. [ Links ]
22. Leon-Quinto T, Jones J, Skoudy A, Burcin M, Soria B. In vitro directed differentiation of mouse embryonic stem cells into insulin-producing cells. Diabetologia 2004; 47: 1442-1451. [ Links ]
23. Damour KA, Bang AG, Eliazer S, Kelly OG, Aqulnick AD, Smart NG et al. Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat Biotechnol 2006; 24: 1392-1401. [ Links ]
24. Poulsom R, Alison MR, Forbes SJ, Wright NA. Adult stem cell plasticity. J Pathol 2002; 197: 441-456. [ Links ]
25. Kojima H, Fujimiya M, Terashima T, Kimura H, Chan L. Extrapancreatic proinsulin/insulin-expressing cells in diabetes mellitus: is history repeating itself? Endocr J 2006; 53: 715-722. [ Links ]
26. Kojima H, Fujimiya M, Matsumura K, Nakahara T, Hara M, Chan L. Extrapancreatic insulin-producing cells in multiple organs in diabetes. Proc Natl Acad Sci USA 2004; 101: 2458-2463. [ Links ]
27. Lanus A, Holz GG, Theise ND, Hussain MA. In vivo derivation of glucose-competent pancreatic endocrine cells from bone marrow without evidence of cell fusion. J Clin Invest 2003; 111: 843-850. [ Links ]
28. Friedenstein AJ, Chailakhyan RK, Latsinik NV, Panasyuk AF, Keiliss-borok IV. Stromal cells responsible for transferring the microenvironment of the hemopoietic tissues. Cloning in vitro and retransplantation in vivo. Transplantation 1974; 17: 331-340. [ Links ]
29. Jiang Y, Henderson D, Blackstad M, Chen A, Miller RF, Verfaille CM. Neuroectodermal differentiation from mouse multipotent adult progenitor cells. Proc Natl Acad Sci USA 2003; 100 Suppl 1: 11854-11860. [ Links ]
30. Tang DQ, Cao LZ, Burkhardt BR, Xia CQ, Litherland SA, Atkinson MA et al. In vivo and in vitro characterization of insulin-producing cells obtained from murine bone marrow. Diabetes 2004; 53: 1721-1732. [ Links ]
31. Moriscot C, De Fraipont F, Richard MJ, marchand M, Savatier P, Bosco D et al. Human bone marrow mesenchymal stem cells can express insulin and key transcription factors of the endocrine pancreas developmental pathway upon genetic and/or microenvironmental manipulation in vitro. Stem Cells 2005; 23: 594-603. [ Links ]
32. Chen G, Han G, Wang J, Wang R, Xu R, Shen B et al. Essential roles of TGF-beta in anti-CD3 antibody therapy: reversal of diabetes in nonobese diabetic mice independent of Foxp3+CD4+ regulatory T cells. J Leukoc Biol. 2008; 83: 280-287. [ Links ]
33. Le Blanc K, Ringdén O. Use of mesenchymal stem cells for the prevention of immune complications of hematopoietic stem cell transplantation. Haematologica 2005; 90: 438. [ Links ]
34. Le Blanc K, Rasmusson I, Sundberg B, Götherström C, Hassan M, Uzunel M et al. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet 2004; 363: 1439-1441. [ Links ]
35. Lazarus HM, Koc ON, Devine SM, Curtin P, Maziarz RT, Holland HK et al. Cotransplantation of HLA-identical sibling culture-expanded mesenchymal stem cells and hematopoietic stem cells in hematologic malignancy patients. Biol Blood Marrow Transplant 2005; 11: 389-398. [ Links ]
36. Kodama S, Kuhtreiber W, Fujimura S, Dale Ea, Faustman DL. Islet regeneration during the reversal of autoimmune diabetes in NOD mice. Science 2003; 302: 1223-1227. [ Links ]
37. Chong S, Normile D. Stem cells. How young Korean researchers helped unearth a scandal. Science 2006; 311: 22-25. [ Links ]
38. Nishio J, Gaglia JL, Turvey SE, Campbell C, Benoist C, Mathis D. Islet recovery and reversal of murine type 1 diabetes in the absence of any infused spleen cell contribution. Science 2006; 311: 1775-1778. [ Links ]
39. Suri A, Calderon B, Esparza TJ, Frederick K, Bittner P, Unanue ER. Immunological reversal of autoimmune diabetes without hematopoietic replacement of beta cells. Science 2006; 311: 1778-1780. [ Links ]
40. Ferber S, Halkin A, Cohen H, Ber I, Einav Y, Goldberg I et al. Pancreatic and duodenal homeobox gene 1 induces expression of insulin genes in liver and ameliorates streptozotocin-induced hyperglycemia. Nat Med 2000; 6: 568-572. [ Links ]
41. Kojima H, Fujimiya M, Matsumura K, Youman P, Imaeda H, Maeda M et al. NeuroD-betacellulin gene therapy induces islet neogenesis in the liver and reverses diabetes in mice. Nat Med 2003; 9: 596-603. [ Links ]
42. Yang L, Li S, Hatch H, Ahrens K, Cornelius JG, Petersen BE et al. In vitro trans-differentiation of adult hepatic stem cells into pancreatic endocrine hormone-producing cells. Proc Natl Acad Sci USA 2002; 99: 8078-8083. [ Links ]
43. Banerjee M, Bhonde RR. Islet generation from intra islet precursor cells of diabetic pancreas: in vitro studies depicting in vivo differentiation. JOP 2003; 4: 137-145. [ Links ]
44. Kodama S, Toyonaga T, Kondo T, Matsumoto K, Tsuruzoe K, Kawashim J et al. Enhanced expression of PDX-1 and Ngn3 by exendin-4 during beta cell regeneration in STZ-treated mice. Biochem Biophys Res Commun 2005; 327: 1170-1178. [ Links ]
45. Suzuki A, Nakauchi H, Taniguchi H. Prospective isolation of multipotent pancreatic progenitors using flow-cytometric cell sorting. Diabetes 2004; 53: 2143-2152. [ Links ]
46. Beattie GM, Rubin JS, Mally MI, Otonkoski T, Hayek A. Regulation of proliferation and differentiation of human fetal pancreatic islet cells by extracellular matrix, hepatocyte growth factor, and cell-cell contact. Diabetes 1996; 45: 1223-1228. [ Links ]
47. Ramiya VK, Maraist M, Arfors KE, Schatz DA, Peck AB, Cornelius JG. Reversal of insulin-dependent diabetes using islets generated in vitro from pancreatic stem cells. Nat Med 2000; 6: 278-282. [ Links ]
48. Ruhnke M, Ungefroren H, Nussler A, Martin F, Brulport M, Schormann W et al. Differentiation of in vitro-modified human peripheral blood monocytes into hepatocyte-like and pancreatic islet-like cells. Gastroenterology 2005; 128: 1774-1786. [ Links ]
49. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006; 126: 663-676. [ Links ]
50. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007; 131: 861-872. [ Links ]
51. Hanna J, Wernig M, Markoulaki S, Sun CW, Meissner A, Cassady JP et al. Treatment of sickle cell anemia mouse model with iPS cells generated from autologous skin. Science 2007; 318: 1920-1923. [ Links ]
52. Stadtfeld M, Brennand K, Hochedlinger K. Reprogramming of pancreatic beta cells into induced pluripotent stem cells. Curr Biol. 2008; 18: 890-894. [ Links ]
53. Tateishi K, He J, Taranova O, Liang G, DAlessio AC, Zhang Y. Generation of insulin-secreting islet-like clusters from human skin fibroblasts. J Biol Chem 2008. [http://www.jbc.org/cgi/doi/10.1074/jbc.M806597200] [ Links ]
54. Zhou Q, Brown J, Kanarek A, Rajagopal J, Melton DA. In vivo reprogramming of adult pancreatic exocrine cells to beta-cells. Nature. 2008; 455: 627-632. [ Links ]
![]() Dirección para correspondencia:
Dirección para correspondencia:
Miguel Barajas
Área de Terapia Celular
Clínica Universitaria
Avda. Pío XII, 36
31008 Pamplona
Tfno. 948255400-ext. 5164
E-mail: mbarajas@unav.es

