










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkInternational Microbiology
versión impresa ISSN 1139-6709
INT. MICROBIOL. vol.8 no.4 dic. 2005
| RESEARCH ARTICLE | |||
|
| |||
| Statistical optimization of a culture medium for biomass and poly(3-hydroxybutyrate) production by a recombinant Escherichia coli strain using agroindustrial byproducts
Summary. A statistically based Plackett-Burman screening design identified milk whey and corn steep liquor concentrations as well as ionic strength (based on phosphate buffer concentration) as the three main independent components of the culture medium that significantly (p < 0.05) influenced biomass and poly(3-hydroxybutyrate) (PHB) production in recombinant cells of Escherichia coli. This strain carries a plasmid encoding phb genes from a natural isolate of Azotobacter sp. Response surface methodology, using a central composite rotatable design, demonstrated that the optimal concentrations of the three components, defined as those yielding maximal biomass and PHB production in shaken flasks, were 37.96 g deproteinated milk whey powder/l, 29.39 g corn steep liquor/l, and 23.76 g phosphates/l (r2 = 0.957). The model was validated by culturing the recombinant cells in medium containing these optimal concentrations, which yielded 9.41 g biomass/l and 6.12 g PHB/l in the culture broth. Similar amounts of PHB were obtained following batch fermentations in a bioreactor. These results show that PHB can be produced efficiently by culturing the recombinant strain in medium containing cheap carbon and nitrogen sources. [Int Microbiol 2005; 8(4):243-250] Key words: recombinant Escherichia coli · biomass · poly(3-hydroxybutyrate) production · agroindustrial by-products · medium optimization | ||
| |||
Optimización estadística de un medio para producción de biomasa y poli(3-hidroxibutirato) por una cepa recombinante de Escherichia coli utilizando subproductos agroindustriales Resumen. Un diseño estadístico de selección de Plackett-Burman identificó las concentraciones de suero de leche y de macerado de maíz, así como la fuerza iónica (dada por la concentración del tampón de fosfatos), como tres variables principales e independientes del medio de cultivo que, de forma significativa (p < 0,05), influían en el crecimiento y la acumulación de biomasa y poli(3-hidroxibutirato) (PHB) en células recombinantes de Escherichia coli. Esta cepa lleva un plásmido que codifica los genes phb provenientes de un aislado natural de Azotobacter sp. Aplicando la metodología de superficies de respuesta, mediante un diseño central compuesto direccionable, se demostró que los valores óptimos de las variables del proceso para la máxima producción de biomasa y de PHB eran: 37,96 g/l de suero de leche desproteinizado en polvo, 29,39 g/l de macerado de maíz y 23,76 g/l de fosfatos (r2 = 0,957). En la validación del modelo, realizada utilizando los valores óptimos, se obtuvieron unas concentraciones de biomasa de 9,41 g/l y de PHB de 6,12 g/l en el medio. En los ensayos en lote en biorreactor se obtuvieron contenidos semejantes de PHB. Los resultados demostraron que el biopolímero puede producirse eficazmente con esta cepa recombinante a partir de fuentes de carbono y nitrógeno de bajo costo. [Int Microbiol 2005; 8(4):243-250] Palabras clave: Escherichia coli recombinante · biomasa · producción de poli(3-hidroxibutirato) · subproductos agroindustriales · optimización del medio de cultivo | Otimização statística do meio pella produção de biomassa e poli(3-hidroxibutirato) por una linhagem recombinante de Escherichia coli usando subproduitos agroindustrais Resumo. Um desenho de seleção, baseado no método estatístico Plackett-Burman identificou as concentrações do soro de queijo e de licor de milho bem como a força iônica (baseada em concentração do tampão fosfatos) as principais variáveis independentes do meio de cultura com significância (p < 0,05), os quais influíam na produção de biomassas e poli(3-hidroxibutirato) (PHB) em células recombinantes de Escherichia coli. Este linhagem leva un plasmídeo que codifica os genes phb de uma linhagem selvagem de Azotobacter sp. A metodologia de superficies de resposta, usando o desenho rotável compósito central, demonstrou que os valores ótimos das variáveis do processo para a máxima produção de biomassa e PHB foram 37,96 g/l de soro de queijo deproteinado, 23,76 g/l de fosfato (r2 = 0,957) e 29,39 g/l de licor de milho. Na validação do modelo, feita usando esses valores ótimos, observaramse unas concentrações de biomassa de 9,41 g/l biomassa e 6,12 g/l de concentração de PHB no caldo de cultivo. Culturos no bioreactor mostraram resultados semelhantes. Os resultados demostraram que o biopolímero pode ser produzido eficientemente com esta linhagem recombinante a partir de fontes econômicas de carbono e nitrogênio. [Int Microbiol 2005; 8(4):243-250] Palavras chave: Escherichia coli recombinante · biomassa · produção de poli(3-hidroxibutirato) · subproduitos agroindustrais · otimização do meio de cultivo |
Introduction
Polyhydroxyalkanoates (PHAs) are a group of polyesters produced by a large number of bacteria that naturally accumulate them in intracellular granules in response to environmental stress and nutrient imbalance [2,10,16]. These thermoplastic polymers have been the subject of great interest since their discovery due to their total biodegradability and the potential to produce them from renewable carbon and nitrogen sources [4,8,11]. Poly(3-hydroxybutyrate) (PHB) is the most widely studied and best characterized PHA. The genes responsible for PHB biosynthesis in a soil isolate of Azotobacter sp. have been cloned and expressed in Escherichia coli [14]. A recombinant strain containing a plasmid that allows expression of these genes was used in the present study. This strain is able to accumulate PHB using lactose as substrate.
In spite of the advantages of PHAs compared with petroleum-derived plastics, their use is currently limited due to their high production costs. Thus, optimization of the fermentation medium is included in the strategic analysis of the viability of this biotechnological process. Optimization of any bioprocess can be conducted either by changing one factor at a time or by varying several factors at the same time and examining their effects and interactions using statistical analysis. The statistical design of experiments is an organized approach that yields more reliable information per experiment than unplanned approaches [7,9,13]. Statistical data analysis enables visualization of the interactions among several experimental variables, leading to the prediction of data in areas not directly covered by experimentation. To date, this sequential statistical methodology has been applied to the screening and optimization of PHB production only for a natural producer microorganism, Alcaligenes latus, grown in a synthetic culture medium [6].
Milk whey is the major by-product of cheese manufacturing, representing 80-90% of the volume of milk transformed. It contains approximately 4.5% (w/v) lactose, 0.8% (w/v) protein, 1% (w/v) salts, and 0.1-0.8% (w/v) lactic acid. Disposal of milk whey is difficult due to its high biological oxygen demand (40 g/l) [1]. Although a fraction of this by-product is used for commercial purposes, such as food ingredients and animal feed, in several countries whey must be treated as a pollutant and disposed of at considerable cost.
Economic evaluation of the PHB production process has suggested that the major contributor to the overall cost is the cost of carbon substrate (up to 50%) [4]. Thus, it is desirable to produce PHB from cheap carbon sources or even from a waste product, such as milk whey. Moreover, there is also a need for low-cost nitrogen sources to reduce PHB production costs.
The aim of this study was to define both the major components of PHB production medium and their optimal concentrations for maximal biomass and biopolymer production. To this end, a new recombinant E. coli strain, and agroindustrial by-products as main carbon and nitrogen substrates were used. In addition, systematic and statistical optimization were carried out using a Plackett-Burman screening design and response surface methodology.
Materials and methods
Bacteria, plasmid and growth conditions. The following Escherichia coli strains were used: CGSC 5040 [F- fadE62 lacI60 tyrT58(AS) fabB5 mel-1(supF58)] (E. coli Genetic Stock Center, New Haven, CT, USA) and CGSC 5040(pK24). Plasmid pJP24 harbors the phbBAC synthesis genes [14] cloned into the expression vector pQE3 (Qiagen, Germany). The strains were grown in a minimal basal medium (MM) containing (in g/l): Na2HPO4 6.0, KH2PO4 3.0, NaCl 0.5, MgSO4·7H2O 0.2, thiamine-HCl 0.005, and ampicillin 0.1, (pH 7.20); supplemented with powdered low fat and deproteinated milk whey [68% (w/w) lactose] and corn steep liquor (CSL) [4.5% (w/w) amino nitrogen] as indicated. The ionic strength (IS) of the culture broth was based on the concentrations of the phosphate buffer components Na2HPO4 and KH2PO4 added [6], keeping the same ratio as in the MM. Erlenmeyer flasks (250 ml) were inoculated with 50 ml MM containing an overnight culture of E. coli CGSC 5040(pK24) (grown in MM containing 30 g lactose/l and 20 g CSL/l) at a concentration of 0.01 g cell dry weight/l. The flasks were set up at least in duplicate and incubated at 37°C in an orbital shaker at 250 rpm rotational speed for 48 h.
For the bioreactor assays, a 5.6-l BioFlo 110 fermenter (New Brunswick Scientific, Edison, NJ, USA) was used. The fermenter was equipped with controllers for pH, temperature, agitation, and dissolved oxygen concentration (DOC). Batch cultures were carried out in 4.2 l MM medium containing deproteinated milk whey, CSL, and phosphates at concentrations determined to be optimal (based on results of the experiments carried out in shaken flasks) for biomass and PHB production. Pre-sterilized (118ºC, 20 min) culture medium was inoculated as described above. Temperature was maintained at 37ºC and pH was controlled at 7.20 ± 0.03 by automatic addition of 5 N NaOH. The agitation speed automatically varied at a fixed air flow rate to maintain the DOC at 40% of air saturation during fermentation. To control foam formation, 30 µl antifoam/l (Antifoam289, Sigma, Saint Louis, MO, USA)/l, was added at the beginning of the run. Samples (15 ml) for off-line biomass and PHB determinations were withdrawn every 4 h until the end of the fermentation (48 h).
Cell dry mass and PHB determination. Ten-ml culture samples were centrifuged at 10,000 × g for 10 min at 4°C and cell dry weight (CDW) was gravimetrically determined using the pellet fraction. Cell pellets were washed twice with deionized water, lyophilized, and weighed. Cell concentration was expressed as g CDW/l. PHB was extracted by alkaline treatment with 0.2 N NaOH as described previously [5] and determined gravimetrically. PHB content is defined as the ratio 100 × (g PHB/g CDW).
Experimental designs and statistical analysis. As a first step in the optimization of cell growth conditions and PHB production in shaken flasks, a Plackett-Burman screening design (PBSD) [15] was used to analyze the main medium constituents influencing CDW and PHB production. [15]. The PBSD was set up for three factors, with two coded levels (-1 and +1) and a center point, and was run to evaluate the linear effects of milk whey concentration (X1), which varied from 7.35 to 44.12 g/l; IS (X2), from 2 to 20 g/l phosphates; and CSL concentration (X3), from 2.23 to 35.68 g/l. The results were fitted with a first-order model.
A Box-Wilson central composite design (CCD) [3] with five settings for each of the three factors levels (milk whey, CSL, and IS) was used to evaluate the quadratic effects and two-way interactions among these variables. The concentrations of other media components were kept constant as in MM throughout this investigation. The value of α = 1.682 was selected to make the design rotatable. In both PBSD and CCD, the variable levels Xi were coded as xi according to Eq. 1:
![]()
where xi and Xi are the dimensionless (codified) value and the actual value of an independent variable respectively, X0 is the real value of an independent variable at center point, and ΔXi is the step change.
In CCD, milk whey (X1) had a lower limit of 14.71 g/l and an upper limit of 44.12 g/l, and IS (X2) was varied between 7 and 27 g/l phosphates. The lower and upper limits of CSL (X3) were 9.61 and 36.03 g/l, respectively. Response surface methodology was used to analyze this experimental design. The second-degree model used to fit the response to the independent variables is shown in Eq. 2:
![]()
where Y is the predicted response, xixj are the input variables that influence the response variable Y, β0 is the intercept, βi is the i th lineal coefficient, βii is the ith quadratic coefficient, and βij is the ijth interaction coefficient.
All experimental designs were done at least three times. Statistical and numerical analyses were carried out by means of the analysis of variance (ANOVA) and multiple regression, using the software Essential Regression v. 2.205, Mathematica v. 3.0, and Origin v. 6.0.
Results
Preliminary results showed that incubation temperature (28-42°C), MgSO4·7H2O concentration (1-10 mM), trace elements solution [12] (1-20 ml/l), and inoculum size (0.01-10 g CDW/l) did not significantly affect cell growth and PHB production in MM supplemented with 30 g milk whey/l and 20 g CSL/l. However, levels of milk whey, CSL, and yeast extract (as nitrogen and vitamin sources) as well as increased IS, which have been reported to have an important effect on PHB accumulation [6], were significant factors for both CDW and PHB production (p < 0.05). As yeast extract increased PHB production only at concentrations higher than 5 g/l, its influence on polymer accumulation was not analyzed further, since it is a high-cost substrate, not suitable for any bioprocess aiming to reduce production costs. Additionally, whereas initial medium pH (between 5.5 and 8.5) influenced CDW production, its effect on PHB accumulation was negligible, so that pH was maintained at 7.20.
Thus, subsequent experiments focused on the influence of whey, IS, and CSL levels in MM on response-dependent variables, as measured using a PBSD. Table 1 shows the coded levels and actual values of those variables and the responses measured.

The ANOVA demonstrated that the three parameters selected were significant at p < 0.05 (for both PHB and CDW production), and Fischer"s F test demonstrated that the model applied was significant (details not shown).
The equations obtained for CDW and PHB production were:
CDW (g/l) = 5.729 + 0.389 x1 + 0.437 x2 + 0.284 x3
PHB (g/l) = 1.874 + 0.277 x1 + 0.188 x2 + 0.223 x3
where x1, x2 and x3 represent the codified levels of milk whey, IS, and CSL, respectively.
Based on these results, plots for the responses measured as a function of two variables were constructed. Figure 1 illustrates the positive effect of whey, CSL, and IS on PHB production.
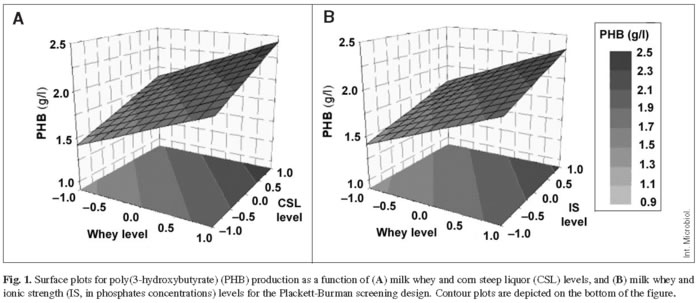
Additionally, to investigate possible interactions among the three variables tested on the responses under study, a two-level fractional factorial design was carried out with the same independent variables and the same range of variation as used for the PBSD. The results of this design were similar to those obtained for the PBSD with respect to the degree of significance (p values) of the variables for CDW and PHB production, and no significant interactions were found (data not shown).
In a second step, the optimal levels of the three significant variables identified in the initial screening experiments were defined in order to obtain maximal CDW and PHB production. Therefore, CCD was carried out to optimize the levels of milk whey, IS, and CSL in the fermentation broth. Table 2 presents the actual and coded values of the three variables in the experimental design, as well as the responses corresponding to CDW and PHB production. Data obtained were analyzed by multiple regression (Table 3). The experimental results of the CCD were fitted and explained with a second-order polynomial function (Eq. 2).


Table 4 shows the results of ANOVA. The lack of fit of the regression model was not significant, and Fischer"s F test demonstrated a high significance (p < 0.05) for the regression. The goodness of fit of the model was checked by the determination coefficient (r2). In this case, the value of the coefficient (r2 = 0.927 for CDW production) indicated that only 7.3% of the total variations are not explained by the model. For PHB production, r2 was 0.957, indicating that only 4.3% of the variations in PHB concentration are not explained by the model. In both cases, the values of the adjusted determination coefficient (adjusted r2 = 0.845 and 0.908 for CDW and PHB production, respectively) indicated a high significance of the model [7]. At the same time, a relatively lower value of the coefficient of variation (CV = 3.78 and 10.15 for CDW and PHB production, respectively) indicated an improved precision and reliability of the experiments [13]. The significance of each coefficient was determined by student"s t test and P values, listed in Table 3, as well as regression coefficients and standard errors. Judging by the regression coefficients and t values [7], it can be concluded that both CDW and PHB production are determined primarily by the linear and quadratic terms of the model, and that no significant interaction exists between any two of the three factors. This conclusion confirms the results previously obtained in the Plackett-Burman and fractional factorial screening designs.

Further proof of the high significance of the model obtained for PHB production is the plot representing predicted versus experimental PHB yields (Fig. 2). Since the plot is very close to y = x (r2 = 0.984), it can be assumed that the prediction of experimental data by the model applied is highly satisfactory.

We then sought to gain additional insight into the optimization of PHB production by E. coli CGSC 5040(pK24), as the polymer is of biotechnological interest. Response surface plots were obtained as a function of two factors at a time, maintaining all other factors at fixed levels. PHB production for different concentrations of the components can be predicted from these plots (Fig. 3). The maximum predicted production of PHB is indicated by the surface confined in the response surface diagram.

Validation of the model and calculation of the regression equation were based on 37.96 g whey/l, 23.76 g phosphates/l, and 29.39 g CSL/l in four independent incubation experiments. The predicted responses were 9.02 g CDW/l and 5.89 g PHB/l, and the actual responses were 9.41 ± 1.35 g CDW/l and 6.12 ± 1.16 g PHB/l, corresponding to a PHB content of 65.03%. The results of the validation experiments proved the applicability of our model.
Based on the results from the flasks cultures, batch cultures of E. coli CGSC 5040(pK24) were set up in a bioreactor. The time course of the fermentation is shown in Fig. 4. PHB production started at about 8 h of fermentation, coinciding with the onset of the stationary growth phase; and then increased linearly up to 48 h of culture. At the end of the fermentation, CDW, PHB concentrations, and PHB content had reached 47.31 g/l, 29.68 g/l, and 62.74%, respectively, implying a volumetric productivity of 0.62 g PHB/(l·h).

Discussion
Biomass and PHB productions were higher in the batch fermentation than in the shaken flasks, resulting in a higher volumetric productivity; however, similar PHB contents were obtained. This indicated that the optimized concentrations for milk whey, CSL, and phosphates for PHB production determined in shaken-flask cultures can be used for PHB production in an aerated bioreactor. In order to improve CDW and PHB productivity, we are currently running fermentations using a fed-batch strategy to feed milk whey and thus avoid the high concentration of acetic acid (ca. 2 g/l) found in the batch cultures, which results in impaired growth (data not shown).
Furthermore, the low-fat and deproteinated milk whey resulted in variations in lactose content of about ± 1.0% for the different batches tested, while for CSL, the variation in amino nitrogen content was about ± 0.5% (data supplied by the manufacturers). Thus, it seems that these agroindustrial by-products can be efficiently used in creating an optimized medium for PHB and CDW production.
In this study, optimal operating conditions to obtain both high biomass and PHB production were determined experimentally by using a response surface methodology. For PHB production, the validity of the model was proved by fitting the values of the variables into a model equation and by actually carrying out the experiments using the optimal concentrations of the three components, both in shaken flasks and in a bioreactor. The statistical strategy as a whole proved to be adequate for the design and optimization of this bioprocess. Moreover, response surface methodology analysis allowed fine-tuning of the nutritional variables in the fermentation medium.
Additionally, we have demonstrated that environmentally friendly PHB polymer can be produced from environmentally polluting whey and a commonly used industrial nitrogen source. The production medium developed in this work may provide an industrially attractive solution to whey disposal and utilization problems, allowing the economical production of high-value biodegradable polyester at the same time. Furthermore, the recombinant strain employed can produce the biopolymer using two agroindustrial by-products as main components of the culture broth, reducing the production costs associated with substrate election, especially in countries where agroindustries have profound impact on the gross domestic product. Additionally, the process used for PHB extraction (alkaline digestion) is not only beneficial from an economical point of view, but also from the point of view of effluent treatment, because the wastes generated in this process are easier to eliminate than those produced in traditional PHA extraction methods (e.g., treatment with chloroform, acetone, detergents, or hypochlorite).
Finally, our results suggest that E. coli CGSC 5040(pK24) is a good candidate for the production of this biopolymer by fermentation, since in medium containing cheap and local sources of carbon and nitrogen as the main substrates, PHB accumulation in minibioreactors, shaken flasks, and a laboratory-scale bioreactor reached up to 65% of the CDW.
Acknowledgements. This research was supported by grants UBACyT X134 and X324, and CONICET PIP 1068. We wish to thank Germán Mauro (Productos de Maíz S. A., Argentina) for the corn steep liquor samples and Dr. Jorge Chirife for the milk whey samples. We are indebted to Dr. Patricia Cerrutti for critically reviewing of the manuscript and Dr. Sandra Cordo for the Portuguese translation. PIN has a graduate student fellowship from CONICET. MJP, BSM and MAG are CONICET researchers.
References
1. Ahn WS, Park SJ, Lee SY (2000) Production of poly(3-hydroxybutyrate) by fed-batch culture of recombinant Escherichia coli with a highly concentrated whey solution. Appl Environ Microbiol 66:3624-3627 [ Links ]
2. Anderson AJ, Dawes EA (1990) Occurrence, metabolism, metabolic role, and industrial uses of bacterial polyhydroxyalkanoates. Microbiol Rev 54:450-472 [ Links ]
3. Box GEP, Wilson KB (1951) On the experimental attainment of optimum conditions. J Roy Statis Soc Ser B 13:1-45 [ Links ]
4. Choi J-I, Lee SY (1997) Process analysis and economic evaluation for poly(3-hydroxybutyrate) production by fermentation. Bioprocess Eng 17:335-342 [ Links ]
5. Choi J-I, Lee SY (1999) Efficient and economical recovery of poly(3-hydroxybutyrate) from recombinant Escherichia coli by simple digestion with chemicals. Biotechnol Bioeng 62:546-553 [ Links ]
6. Grothe E, Moo-Young M, Chisti Y (1999) Fermentation optimization for the production of poly(b-hydroxybutyric acid) microbial thermoplastic. Enzyme Microb Technol 25:132-141 [ Links ]
7. Haaland P (1987) Experimental design in biotechnology. Marcel Dekker, New York, NY. [ Links ]
8. Hänggi UJ (1995) Requirements on bacterial polyesters as future substitute for conventional plastics for consumer goods. FEMS Microbiol Rev 16:213-220 [ Links ]
9. Kalil SJ, Maugeri F, Rodrigues MI (2000) Response surface analysis and simulation as a tool for bioprocess design and optimization. Process Biochem 35:539-550 [ Links ]
10. Khanna S, Srivastava AK (2005) Recent advances in microbial polyhydroxyalkanoates. Process Biochem 40:607-619 [ Links ]
11. Kim BS (2000) Production of poly(3-hydroxybutyrate) from inexpensive substrates. Enzyme Microb Technol 27:774-777 [ Links ]
12. Lee SY, Chang HN (1993) High cell density cultivation of Escherichia coli W using sucrose as a carbon source. Biotechnol Lett 15:971-974 [ Links ]
13. Montgomery DC (2001) Design and analysis of experiments. John Wiley & Sons, New York [ Links ]
14. Pettinari MJ, Vázquez GJ, Silberschmidt D, Rehm B, Steinbüchel A, Méndez BS (2001) Poly(3-hydroxybutyrate) synthesis genes in Azotobacter sp. strain FA8. Appl Environ Microbiol 67:5331-5334 [ Links ]
15. Plackett RL, Burman JP (1946) The design of optimum multifactorial experiments. Biometrika 33:305-325 [ Links ]
16. Vázquez GJ, Pettinari MJ, Méndez BS (2003) Evidence of an association between poly(3-hydroxybutyrate) accumulation and phosphotransbutyrylase expression in Bacillus megaterium. Int Microbiol 6:127-129 [ Links ]

