










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkInternational Microbiology
versión impresa ISSN 1139-6709
INT. MICROBIOL. vol.8 no.4 dic. 2005
| RESEARCH ARTICLE | |||
|
| |||
| Phenotypic, genotypic, and phylogenetic discrepancies to differenciate Aeromonas salmonicida from Aeromonas bestiarum
Summary. The taxonomy of the "Aeromonas hydrophila" complex (comprising the species A. hydrophila, A. bestiarum, A. salmonicida, and A. popoffii) has been controversial, particularly the relationship between the two relevant fish pathogens A. salmonicida and A. bestiarum. In fact, none of the biochemical tests evaluated in the present study were able to separate these two species. One hundred and sixteen strains belonging to the four species of this complex were identified by 16S rDNA restriction fragment length polymorphism (RFLP). Sequencing of the 16S rDNA and cluster analysis of the 16S-23S intergenic spacer region (ISR)-RFLP in selected strains of A. salmonicida and A. bestiarum indicated that the two species may share extremely conserved ribosomal operons and demonstrated that, due to an extremely high degree of sequence conservation, 16S rDNA cannot be used to differentiate these two closely related species. Moreover, DNA-DNA hybridization similarity between the type strains of A. salmonicida subsp. salmonicida and A. bestiarum was 75.6 %, suggesting that they may represent a single taxon. However, a clear phylogenetic divergence between A. salmonicida and A. bestiarum was ascertained from an analysis based on gyrB and rpoD gene sequences, which provided evidence of a lack of congruence of the results obtained from 16S rDNA, 16S-23S ISR-RFLP, DNA-DNA pairing, and biochemical profiles. [Int Microbiol 2005; 8(4):259-269] Key words: Aeromonas spp. · nucleic acids techniques · gene gyrB · gene rpoD | ||
| |||
Discrepancias fenotípicas, genotípicas y filogenéticas para diferenciar Aeromonas salmonicida de Aeromonas bestiarum Resumen. La taxonomía del complejo "Aeromonas hydrophila" (que comprende las especies A. hydrophila, A. bestiarum, A. salmonicida, y A. popoffii) ha sido controvertida, en particular las relaciones entre los dos patógenos de peces, A. salmonicida y A. bestiarum. De hecho, de las pruebas bioquímicas evaluadas en el presente estudio, ninguna fue capaz de separar estas dos especies. Ciento dieciséis cepas pertenecientes a las cuatro especies de este complejo se identificaron mediante el análisis del polimorfismo de la longitud de los fragmentos de restricción (RFLP) del 16S rDNA. La secuenciación del 16S rDNA y el análisis de grupos de RFLP de la región espaciadora intergénica (ISR) 16S-23S en cepas seleccionadas de A. salmonicida y A. bestiarum indicaron que las dos especies podrían compartir operones ribosómicos extremadamente conservados y demostraron que, debido a su elevado grado de conservación de secuencia, el 16S rDNA no puede utilizarse para diferenciar estas dos especies de relación tan estrecha. Además, la similitud de hibridación DNA-DNA entre las cepas tipo de A. salmonicida subsp. salmonicida y de A. bestiarum era del 75,6 %, lo que sugiere que pueden ser un único taxón. Sin embargo, el análisis simultáneo de las secuencias de los genes gyrB y rpoD puso de manifiesto una marcada divergencia filogenética entre A. salmonicida y A. bestiarum, lo cual aporta pruebas de la falta de congruencia de los resultados de 16S rDNA, ISR-RFLP, 16S-23S, apareamiento DNA-DNA y perfiles bioquímicos. [Int Microbiol 2005; 8(4):259-269] Palabras clave: Aeromonas spp. · técnicas de ácidos nucleicos · gen gyrB · gen rpoD | Discrepancias fenotípicas, genotípicas e filogenéticas para diferenciar Aeromonas salmonicida de Aeromonas bestiarum Resumo. A taxonomia do complexo "Aeromonas hydrophila" (que compreende as espécies A. hydrophila, A. bestiarum, A. salmonicida, e A. popoffii) tem sido controvérsia, em particular a relacionada com as duas espécies patogênicas em peixes, A. salmonicida e A. bestiarum. De facto, com as provas bioquímicas efectuadas no presente estudo, observou-se que nenhuma foi capaz para separar estas duas espécies. Cento e dezasseis estirpes pertencentes às quatro espécies deste complexo identificaram-se por a análise do polimorfismo no tamanho dos fragmentos de restrição (RFLP) do 16S rDNA. A sequênciação do 16S rDNA e a análise dos grupos de RFLP da região espaciadora intergênica (ISR) 16S-23S em estirpes seleccionadas de A. salmonicida e de A. bestiarum indicaram que as duas espécies poderão partilhar operões ribossómicos extremamente conservados e demonstram que, devido ao seu elevado grau de conservação da sequência, o 16S rDNA não se pode utilizar para diferenciar estas duas espécies de relação tão estreita. Por outro lado, a semelhança de hibridação DNA-DNA entre estirpes tipo de A. salmonicida subsp. salmonicida e de A. bestiarum foi de 75,6% sugerindo que podem representar um único taxon. No entanto, a análise simultânea das sequências dos genes gyrB e rpoD apresenta uma notável divergência filogenética entre A. salmonicida e A. bestiarum, o qual evidencia uma falta de congruência no que diz respeito aos resultados de 16S rDNA, 16S-23S ISR-RFLP, emparelhamento DNA-DNA e perfis bioquímicos. [Int Microbiol 2005; 8(4):259-269] Palavras chave: Aeromonas spp. · técnicas de ácidos nucleicos · gene gyrB · gene rpoD |
Introduction
Aeromonads are autochthonous to aquatic environments worldwide and have been strongly implicated in the etiology of a variety of fish and human diseases. Particularly relevant to aquaculture is the species A. salmonicida, the causative agent of fish furunculosis. The taxonomy of the genus Aeromonas is complex and some of its species cannot be easily differentiated [17]. DNA-DNA reassociation studies carried out by Popoff in the early 1980s [44,45] identified three different hybridization groups (HGs) within the phenotypic "A. hydrophila" complex: HG1, represented by A. hydrophila; HG2, initially represented by a group of unclassified strains; and HG3, A. salmonicida. The latter species now includes non-motile strains isolated from fish divided into four psycrophilic subspecies: A. salmonicida subsp. salmonicida [18], A. salmonicida subsp. achromogenes [47], A. salmonicida subsp. masoucida [32,33], A. salmonicida subsp. smithia [7], and the mesophilic subspecies A. salmonicida subsp. pectinolytica isolated from water [43]. In addition, A. salmonicida includes motile mesophilic strains isolated from diverse sources, confusingly referred to as "A. hydrophila" (HG3) and classically represented by the reference strain Popoff 316 [5]. The complexity of A. salmonicida taxonomy is reflected by the number of articles describing strains that cannot be included in any of the described subspecies due to the presence of atypical biochemical and genetic characteristics [6,7,22,35,46,50,52]. The unnamed HG2 was first called A. salmonicida subsp. bestiarum [10,30], but later, although it showed high (71%) DNA-DNA reassociation with "A. hydrophila" (HG3) and even higher (78%) with A. hydrophila (HG1), it was designated as a different species, A. bestiarum [3]. In addition, the type strain of A. salmonicida was not hybridized with A. bestiarum in the definition of the latter species. By contrast, the 16S rRNA gene sequence of the type strain of A. bestiarum is identical to that of A. salmonicida subsp. achromogenes and A. salmonicida subsp. masoucida and only shows two nucleotide differences from that of A. salmonicida subsp. salmonicida [36,38]. By using the RFLPs of the 16S rRNA gene, we obtained species-specific patterns for all of the type strains of the genus [9,17]. Recently, however, we have isolated numerous strains (referred to here as A. bestiarum/A. salmonicida) displaying a pattern that is a mixture of those shown by A. bestiarum and A. salmonicida strains. The present report attempts to determine whether there is genetic or phenotypic distinction between strains of these two specie as well as among strains of other species that integrate the "A. hydrophila" complex, including the recently described species A. popoffii.
Materials and methods
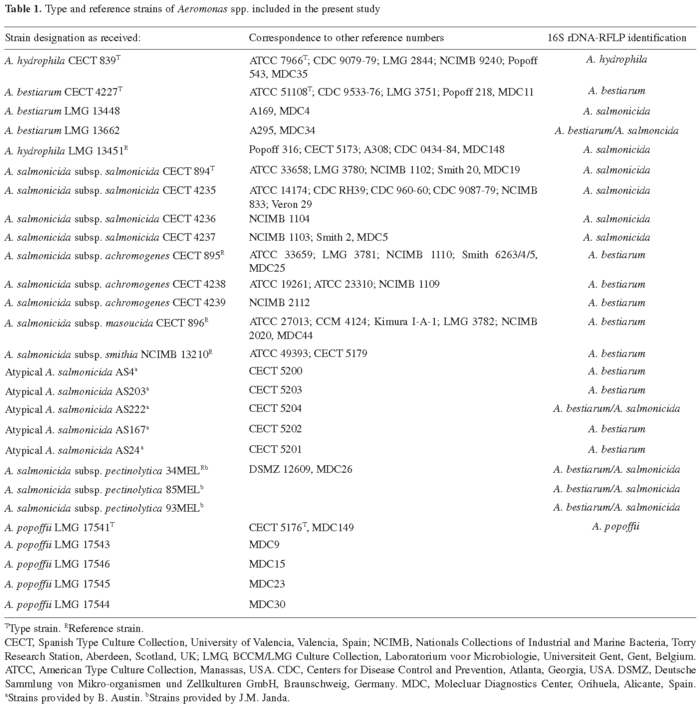
Bacterial strains and culture conditions. A total of 116 strains identified by the 16S rDNA-RFLP technique [9,17] were included in the study. The isolates were grown on TSA (Trypticase Soy Agar, Difco; Barcelona, Spain) at 30oC, with the exception of those belonging to A. salmonicida subsp. salmonicida, A. salmonicida subsp. masoucida, A. salmonicida subsp. achromogenes, A. salmonicida subsp. smithia and "atypical" A. salmonicida isolates, all of which were grown at room temperature (20-22). Ninety-three of isolates (18 A. hydrophila, 7 A. bestiarum, 26 A. bestiarum/A. salmonicida, 18 A. salmonicida and 24 A. popoffii) were predominantly environmental isolates from our collection, 22 were type and reference strains, and five were "atypical" A. salmonicida strains supplied by B. Austin (Heriot-Watt University, Edinburgh, Scotland) (Table 1).
Biochemical studies. The following biochemical tests were done in all strains according to conventional protocols: esculin hydrolysis, lysine decarboxylase, citrate utilization, motility, indole production, N-acetyl-D-glucosamine utilization, gluconate oxidation and acid production from rhamnose, sorbitol, lactose, D-sucrose, and salicin. DL-lactate utilization was tested according to the method of Janda et al. [29], urocanic acid according to the method of Hänninen [19] and elastase according to the method of Hasan et al. [20]. Test samples were incubated under the same conditions as used for bacterial growth. Readings were made every day for 3 days, and up to 5 days for the elastase test. All tests were carried out in duplicate, and appropriate positive and negative controls were included. Growth temperature (4°C, 20-22°C, 30°C, 37°C, and 44.5°C) and brown diffusible pigment production were evaluated on TSA [6]. Readings were taken every 24 h for up to 5 days for growth and up to 7 days for pigment production.
16S rDNA-RFLP identification. Briefly, the PCR-amplified 16S rRNA gene (1502 bp) was digested using two endonucleases (AluI and MboI) simultaneously, thus enabling the identification of all the species of the genus, with the exception of A. bestiarum, A. salmonicida, A. popoffii, and A. encheleia. To differentiate the latter two, further digestion with NarI, followed by digestion with HaeIII and AlwNI, was necessary, while A. salmonicida and A. bestiarum were distinguished by digestion with NarI followed by PstI [17].
16S rRNA gene sequencing. A fragment of ca. 560 bp of the 16S rRNA gene of the A. bestiarum and A. salmonicida type strains and of a subgroup of 14 isolates characterized as A. bestiarum/A. salmonicida by 16S rDNA-RFLP was sequenced. This fragment contains the characteristic variable positions (1011 and 1018 bp) for A. bestiarum and A. salmonicida [38]. Genomic DNA extraction and PCR amplification were performed as previously described [9,38]. PCR products were purified using Concert Rapid PCR Purification System (Life Technologies; Barcelona, Spain) following the manufacturer"s instructions. Sequencing primers were those described in Martinez-Murcia et al. [37] and sequences were determined using the ThermoSequenase fluorescently labeled primer cycle sequencing kit with 7-deaza-dGTP (Amersham Pharmacia Biotech; Barcelona, Spain) and the ALFexpressII sequencer (Amersham) according to the manufacturer"s indications.
DNA-DNA hybridization. DNA was isolated by chromatography on hydroxyapatite [11] and further submitted to DNA-DNA hybridization at 66oC according to the method of De Ley et al. [13], with some modifications [14,23]. Samples were read using a 2600 spectrophotometer equipped with a 2527-R thermoprogrammer and plotter (Gilford, Oberlin, Ohio, USA). Renaturation rates were computed with the TRANSFER.BAS program [27].
16S-23S ISR-RFLP. Seventy-six of the strains, including a representative number of each species, were analyzed by the 16S-23S intergenic spacer region-restriction fragment length polymorphism (ISR-RFLP) technique. Genomic DNA extraction and PCR amplification were performed as previously described [39,48]. Two double digestions of the 16S-23S ISR were carried out using the HinfI-CfoI and HinfI-TaqI (Gibco BRL, Barcelona, Spain). Digestion products were electrophoresed on a 4% metaphore agarose gel (FMCS Bioproducts Europe, Denmark). Gel images were saved as TIFF files and further analyzed by BioNumerics software, version 1.5 (Applied Maths, Kortrijk, Belgium). Levels of similarity between the profiles were calculated using the band-matching Dice coefficient (SD), and the cluster analysis of similarity matrices was calculated with the unweighted pairgroup method with arithmetic means (UPGMA).
PCR amplification and sequencing of gyrB and rpoD. Fragments of ca. 1100 bp and 820 bp of the gyrB and rpoD genes, respectively, were amplified by PCR and sequenced as previously described [49,54].
Phylogenetic data analysis. The nucleotide sequences of gyrB and rpoD were aligned, treated as a single nucleotide sequence [53], and the evolutionary tree was constructed as previously described [49].
Results
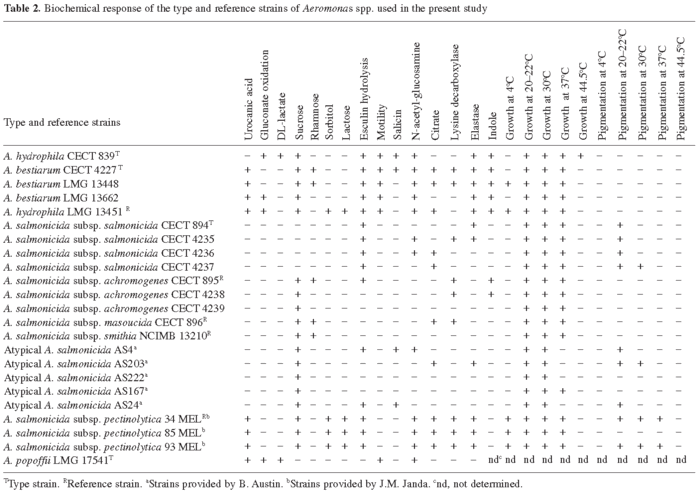
Biochemical studies. Table 2 shows the biochemical responses obtained with the type and reference strains of Aeromonas included in the study. All strains of A. salmonicida subsp. pectinolytica and "A. hydrophila" LMG 13451 (Popoff 316) were able to grow from 4 to 37°C. The remaining A. salmonicida subspecies strains grew from room temperature up to 37°C, and "atypical" A. salmonicida isolates up to 30°C, with the exception of strain AS167, which could grow also at 37°C. Brown diffusible pigment production was observed in two strains of A. salmonicida subsp. pectinolytica, all strains of A. salmonicida subsp. salmonicida, and in three "atypical" isolates, but this response was temperature-dependent. All strains from our collection grew from 20-22°C up to 37°C, while only some grew also at 4°C or at 44.5°C (data not shown). Brown diffusible pigment production was observed in one A. bestiarum isolate, one A. bestiarum/A. salmonicida isolate, and one of the A. salmonicida isolates (data not shown).
Type strains of A. hydrophila and A. bestiarum and the reference strain "A. hydrophila" LMG 13451 (Popoff 316) were motile and indole-positive, while the type and reference strains of the subspecies of A. salmonicida and the "atypical" isolates were nonmotile and indole-negative (with the exception of two A. salmonicida subsp. Achromogenes, which were indole-positive). The 51 strains of A. salmonicida, A. bestiarum/A. salmonicida and A. bestiarum showed variable motility (67, 69, and 86%, respectively) (Table 3) and indole production (86, 75, and 25 %, respectively) (data not shown).
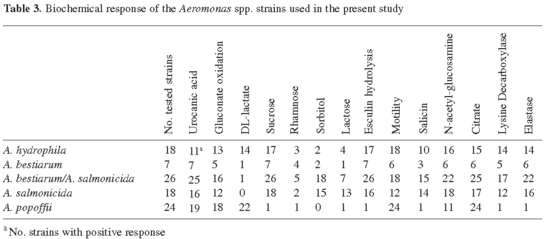
The analysis of DL-lactate utilization allowed separation of 78% of the strains of A. hydrophila from A. bestiarum and A. salmonicida. None of the 14 biochemical tests could unequivocally distinguish A. bestiarum from A. salmonicida, nor from the subspecies of A. salmonicida. However, positive acid production from sorbitol and lactose was more prevalent for strains of A. salmonicida than for those of A. bestiarum. The analysis of DL-lactate utilization, acid production from sucrose, esculin hydrolysis, and elastase allowed discrimination between A. popoffii and the rest of the species of the "A. hydrophila" complex.
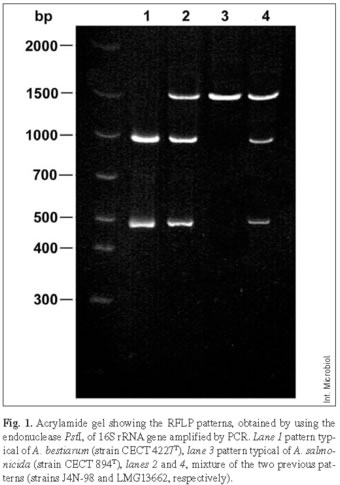
16S rDNA-RFLP and 16S rDNA sequencing. The typical 16S rDNA-RFLP profiles of the type strains of A. bestiarum (fragments of 1005 bp and 497 bp), A. salmonicida (fragment of 1502 bp), and of strains referred to as A. bestiarum/A. salmonicida, with a mixture of these two patterns, are shown in Fig. 1. Variation in band intensity was observed in the latter as: (i) similar intensity of the three bands (i.e., strain J4N-98) and (ii) a relatively higher intensity of the 1502-bp band (i.e., strain LMG 13662).
Three subspecies of A. salmonicida, i.e., achromogenes, masoucida, and smithia, and four "atypical" A. salmonicida isolates yielded the 16S-RFLP pattern of A. bestiarum, while strain "A. hydrophila" LMG 13451 (Popoff 316) showed the pattern of A. salmonicida. Furthermore, of the 26 strains from our collection included in the study, one "atypical" A. salmonicida strain (AS222), three A. salmonicida subsp. pectinolytica strains (34 MEL, 85 MEL and 93 MEL), and the strain LMG 13662 used in the description of the species A. bestiarum [3] showed the mixed A. bestiarum/A. salmonicida 16S rDNA-RFLP pattern (see Table 1).
The sequences of the 16S rDNA gene of the type strains showed the expected nucleotides at the variable positions 1011 and 1018, which correspond to T and A for A. salmonicida, and C and G for A. bestiarum. However, the 14 A. bestiarum/A. salmonicida strains selected for sequencing gave ambiguities suggesting micro-heterogeneity. Visual inspection of the chromatograph showed a double sequencing signal at these positions, although secondary peaks were slightly smaller. Two types of double sequencing peaks were discerned and they were in accordance with the obtained RFLP intensity profiles. When the 16S rDNA-RFLP pattern showed equal intensity of the three bands, sequencing analysis showed C/T and G/A at positions 1011 and 1018, respectively, while T/C and A/G were obtained with higher intensity of the 1502 bp band (Fig. 1 and Table 4).

DNA-DNA hybridization. DNA-DNA hybridization results are shown in Table 5. The strain "A. hydrophila" HG3 (Popoff 316) hybridized 88.3% with the type strain of A. salmonicida (A. salmonicida subsp. salmonicida, CECT 894T) and with a similarity ranging from 79.6 to 83.8% with the reference strains of the other subspecies. In addition, the type strain of A. bestiarum (CECT 4227T) showed 75.6% similarity with the type strain of A. salmonicida (A. salmonicida subsp. salmonicida, CECT 894T) and similarities ranging from 70.5 to 83.5% with reference strains of other subspecies.

16S-23S ISR-RFLP analyses. Using HinfI-CfoI, digestions of the 16S-23S ISR produced five to nine bands per strain, while with HinfI-TaqI, four to 12 bands were obtained; all the bands were smaller than 500 bp. The UPGMA dendogram derived from the analysis of the combined profiles of HinfI-CfoI and HinfI-TaqI is shown in Fig. 2. The two main clusters, one containing the A. popoffii strains and the other the rest of the strains, were separated by a coefficient value of <20%. The latter cluster split into two subclusters, one that encompassed the strains of A. hydrophila (at a similarity level of 52%) and the other containing the strains of A. bestiarum, A. salmonicida, and A. bestiarum/A. salmonicida interspersed (at 62% profile similarity).
gyrB and rpoD phylogenetic analysis. Nucleotide sequences of gyrB and rpoD amplicons were determined from representative strains of these four Aeromonas species. The corresponding sequences of reference strains were previously published [49,54]. Experiments were repeated at least twice from single colonies of original cultures to confirm readings and resolve ambiguities. The derived gyrB sequences comprised between 960 and 1100 nucleotides and the rpoD sequences between 813 and 825 nucleotides. The unrooted phylogenetic tree, constructed by using the combined gyrB-rpoD gene sequences (1770 bp; Fig. 3), showed a clear clustering of these species.
Discussion
Although several approaches have been attempted aimed at clarifying the phylogenetic relationships among the species of the genus [24,36,38], the taxonomy of Aeromonas remains difficult. One of the most evident problems lies in the delineation of the species of the "A. hydrophila" complex, since DNA-DNA reassociation results between A. bestiarum and A. salmonicida [3,4,19] do not provide conclusive information.
An interesting aspect observed in the present study was that A. salmonicida showed mesophilic behavior in being able to grow at 37°C. This species had been traditionally considered psycrophilic, with optimal growth at 22°C-25°C [6,7,32,33,44]. Its response to temperature is identical to that of A. bestiarum [3].
Classical tests used to separate the subspecies of A. salmonicida from each other are motility and indole production [43,44,52]. However, these properties did not prove useful for that purpose in our study. Motility only yielded the expected results for the type and reference strains, whereas it was very variable in other strains. Similarly, indole production was also a highly variable character and therefore inappropriate for subspecies differentiation. Another feature traditionally used to differentiate A. salmonicida subsp. salmonicida is the production of brown diffusible pigment, which is lacking in the other subspecies [7,32,33,40,52]. However, in agreement with Austin et al. [6], the results of this test were temperature-dependent and not specific for A. salmonicida subsp. salmonicida. This was also observed by Dalsgaard et al. [12], who found that A. salmonicida subsp. achromogenes was able to produce a brown diffusible pigment. Urocanic acid utilization was tested for the first time on reference strains of the different subspecies of A. salmonicida and all of them were negative.
Other biochemical tests, such as gluconate oxidation, acid production from lactose and sucrose, esculin hydrolysis, N-acetyl-glucosamine utilization, and elastase production, which have been proposed by different authors to separate subspecies of A. salmonicida [6,7,42,43,52], were unable to discriminate such subspecies-as was demonstrated previously by Noterdaeme et al. [41] using 128 biochemical and physiological characters.
Similar difficulties in the biochemical identification of the A. salmonicida subspecies have been found by other authors, who then had to consider numerous isolates of A. salmonicida as "atypical" [6,7,22,35,50,52]. Although the synonymization of the subspecies of A. salmonicida has been under discussion for several years [7, 8, McCarthy DH, 1978, PhD thesis], no consensus has ever been reached, and even a new subspecies, A. salmonicida subsp. pectinolytica, was proposed recently [43]. Our results demonstrate that strains of the latter subspecies also share ribosomal operons from both A. bestiarum and A. salmonicida.
An extensive revision of the biochemical tests proposed by different authors to differentiate the species of the "A. hydrophila" complex [see Table 6 in the online version of this article] revealed the existence of numerous contradictory data, such as acid production from sorbitol [2], and rhamnose [2,3,19], lactose [1-3,31], urocanic acid, and DL-lactate utilization [3], within each of the species A. hydrophila, A. bestiarum, and A. salmonicida. These tests have previously been considered useful to discriminate these three species [1,3,19,29,31]. Our results mostly agree with those of Abbott et al. [1], even though those authors did not include the type species of A. salmonicida, and the strains that we investigated were from more diverse origins.
In our study, DL-lactate utilization and acid production from sorbitol and lactose were the most useful tests to distinguish A. hydrophila (see Table 3), but these tests cannot be considered unequivocal criteria, in agreement with other authors [31,34]. Acid production from D-sucrose, DL-lactate utilization, and lysine decarboxylase activity were used by Huys et al. [25] to separate A. popoffii from species of the "A. hydrophila" complex. In the present study, the tests useful for this purpose were esculin hydrolisis and acid production from D-sucrose. DL-lactate utilization was useful for the separation of A. popoffii from A. bestiarum and A. salmonicida but not from A. hydrophila. Recently, Abbott et al. [1] indicated that A. popoffii is always elastase negative, while in our study only one strain was clearly positive for this test.
Sequencing of the 16S rRNA gene is considered a robust taxonomic tool and is now widely used in bacterial taxonomy. In Aeromonas, this gene provides also signature regions for the delineation and identification of most species [38], but for some strains of some species the sequence was found to be extremely conserved; for example, A. salmonicida and A. bestiarum type strains differ by only two nucleotides, at positions 1011 and 1018. In the present study, RFLP and/or sequencing of the 16S rDNA of a considerable number of the isolates tested (ca. 60% of strains investigated), including some reference strains, suggested that these isolates may share ribosomal operons from both species, A. bestiarum and A. salmoncida. Differences in the intensity of the 16S rDNA-RFLP bands and of the sequencing signals were consistent and may be related to a difference in the number of rrn operon copies of one species or another among strains. Furthermore, 16S-23S ISR-RFLP analysis showed that, while A. hydrophila and A. popoffii formed two different clusters, strains of A. bestiarum, A. bestiarum/ A. salmonicida, and A. salmonicida clustered together in a single group.
DNA-DNA hybridization remains the standard method for species designation [15,51]. In the present study, we determined the DNA-DNA hybridization values for strains that had not previously been evaluated. The type strain for A. bestiarum showed 75.6% hybridization with the type strain for A. salmonicida (A. salmonicida subsp. salmonicida CECT 894T), which is consistent with difficulties found in separating strains of these two species by DNA-DNA pairing [19]. Ali et al. [3] described A. bestiarum as a new species following only a DNA-DNA hybridization with the reference strain of "A. hydrophila" HG3, named Popoff 316. Discordances between different sets of DNA-DNA hybridization data published by different authors using different techniques, but dealing with the same Aeromonas reference strains, have been reported several times for other species of this genus. For instance, strains of A. allosaccharophila exhibited 0-40% DNA relatedness with A. veronii [16], although later DNA-DNA hybridization studies by Huys et al. [26] indicated a much higher range (78-84%) using the same reference strains, but also inconsistency with previous data obtained by Hickman-Brenner et al. [21]. The most pronounced disagreement so far is that regarding A. encheleia/Aeromonas sp. HG11/A. eucrenophila, reported by Martínez-Murcia [33], by comparing data from two previous studies. As suggested, the measure of DNA sequence homology by DNA-DNA hybridization may be too crude, or at least not fine enough to split highly similar species.
However, phylogenetic analysis of gyrB and rpoD housekeeping genes (see Fig. 3) showed that, although genetically related, the strains of A. bestiarum and A. salmonicida form two clearly separated clusters with considerable phylogenetic divergence, in agreement with previous studies based on AFLP fingerprinting [24]. Substitution rates of rpoD between strains of A. salmonicida and A. bestiarum were double those obtained from the corresponding gyrB sequences, as previously reported [49]. New isolates of these two species were also included in the present phylogenetic analysis and the tree topology (clustering and branch distances) was maintained. This result confirmed that gyrB and rpoD are excellent molecular markers for assessing the phylogeny of closely related Aeromonas species, but also indicated a new lack of congruence with results derived from DNA-DNA hybridization data and 16S rDNA sequencing.
In conclusion, the high DNA homology of A. bestiarum and A. salmonicida, the impossibility of separating the two species by biochemical tests, neither by 16S rDNA nor by 16S-23S ISR-RFLP, could be taken as preliminary evidence that they are a single taxon. However, by analysis of faster molecular chronometers (e.g., rpoD, gyrB), the resulting phylogenies indicated that these two species show a modest amount of divergence, considering the overall phylogenetic depth of the genus Aeromonas. This study confirms the limitations of 16S rDNA sequencing to split Aeromonas species and suggests that DNA-DNA hybridization is not reliable when dealing with closely related species. Finally, the use of conserved housekeeping genes is recommended to identify new Aeromonas isolates.
Acknowledgements. This work was supported by grants FIS 03/1183, from the Spanish Ministry of Health, CIRIT (SGR 1999/00103), Generalitat Valenciana (GV98-21-05), Fundació Ciència i Salut, and a fellowship from the Universitat Rovira i Virgili. We would like to thank the Colección Española de Cultivos Tipo (CECT), the Belgium Co-ordinated Collection of Micro-organisms (LMG), Dr. B. Austin, Dr. A. Demarta, Dr. C.I. Kingombe, and Dr. A. Kozi?ska for kindly providing isolates. We thank Bettina Straübler for performing the DNA-DNA hybridization experiments.
References
1. Abbott SL, Cheung WW, Janda JM (2003) The genus Aeromonas: biochemical characteristics, atypical reactions, and phenotypic identification schemes. J Clin Microbiol 41:2348-2357 [ Links ]
2. Abbott SL, Cheung WKW, Kroske-Bystrom S, Malekzadeh T, Janda JM (1992) Identification of Aeromonas strains to the genospecies level in the clinical laboratory. J Clin Microbiol 30:1262-1266 [ Links ]
3. Ali A, Carnahan AM, Altwegg M, Lüthy-Hottenstein J, Joseph SW (1996) Aeromonas bestiarum sp. nov. (formerly genomospecies DNA group 2 A. hydrophila), a new species isolated from non-human sources. Med Microbiol Lett 5:156-165 [ Links ]
4. Altwegg M (1993) A polyphasic approach to the classification and identification of Aeromonas strains. Med Microbiol Lett 2:200-205 [ Links ]
5. Altwegg M, Steigerwalt AG, Altwegg-Bissig R, Lüthy-Hottenstein J, Brenner DJ (1990) Biochemical identification of Aeromonas genospecies isolated from humans. J Clin Microbiol 28:258-264 [ Links ]
6. Austin B, Austin DA, Dalsgaard I, Gudmundsdóttir BK, Hoie S, Thornton JM, Larsen JL, O"Hici B, Powell R (1998) Characterization of atypical Aeromonas salmonicida by different methods. Syst Appl Microbiol 21:50-64 [ Links ]
7. Austin DA, McIntosh D, Austin B (1989) Taxonomy of fish associated Aeromonas spp., with the description of Aeromonas salmonicida subsp. smithia subsp. nov. Syst Appl Microbiol 11:277-290 [ Links ]
8. Belland RJ, Trust TJ (1988) DNA:DNA reassociation analysis of Aeromonas salmonicida. J Gen Microbiol 134:307-315 [ Links ]
9. Borrell N, Acinas SG, Figueras MJ, Martínez-Murcia AJ (1997) Identification of Aeromonas clinical isolates by restriction fragment length polymorphism of PCR-amplified 16S rRNA genes. J Clin Microbiol 35:1671-1674 [ Links ]
10. Carnahan AM (1993) Update on current identification methods for clinical Aeromonas isolates. Med Microbiol Lett 2:212-216 [ Links ]
11. Cashion P, Holder-Franklin MA, McCully J, Franklin M (1977) A rapid method for base ratio determination of bacterial DNA. Anal Biochem 81:461-466 [ Links ]
12. Dalsgaard I, Gudmundsdóttir BK, Helgason S, Høie S, Thoresen OF, Wichardt UP, Wiklund T (1998) Identification of atypical Aeromonas salmonicida: inter-laboratory evaluation and harmonization of methods. J Appl Microbiol 84:999-1006 [ Links ]
13. De Ley J, Cattoir H, Reynaerts A (1970) The quantitative measurement of DNA hybridization from renaturation rates. Eur J Biochem 12:133-142 [ Links ]
14. Escara JF, Hutton JR (1980) Thermal stability and renaturation of DNA in dimethylsulphoxide solutions: acceleration of renaturation rate. Biopolymers 19:1315-1327 [ Links ]
15. Esteve C (1997) Is AFLP fingerprinting a true alternative to the DNA-DNA pairing method to assess genospecies in the genus Aeromonas? Int J Syst Bacteriol 47:245-246 [ Links ]
16. Esteve C, Gutiérrez MC, Ventosa A (1995) Aeromonas encheleia sp. nov., isolated from European eels. Int J Syst Bacteriol 45:462-466 [ Links ]
17. Figueras MJ, Soler L, Chacón MR, Guarro J, Martínez-Murcia AJ (2000) Extended method for discrimination of Aeromonas spp. by 16S rDNA-RFLP analysis. Int J Syst Evol Microbiol 50:2069-2073 [ Links ]
18. Griffin PJ, Snieszko SF, Friddle SB (1953) Pigment formation by Bacterium salmonicida. J Bacteriol 65:652-659 [ Links ]
19. Hänninen ML (1994) Phenotypic characteristics of the three hybridization groups of Aeromonas hydrophila complex isolated from different sources. J Appl Bacteriol 76:455-462 [ Links ]
20. Hasan JAK, Macaluso P, Carnahan AM, Joseph S (1992) Elastolytic activity among Aeromonas spp. using a modified bilayer plate assay. Diagn Microbiol Infect Dis 15:201-206 [ Links ]
21. Hickman-Brenner FW, MacDonald KL, Steigerwalt AG, Fanning GR, Brenner DJ, Farmer III JJ (1987). Aeromonas veronii, a new ornithine decarboxilase-positive species that may cause diarrhea. J Clin Microbiol 25:900-906 [ Links ]
22. Høie S, Dalsgaard I, Aase IL, Heum M, Thornton JM, Powell R (1999) Polymerase chain reaction (PCR)-based typing analysis of atypical isolates of the fish pathogen Aeromonas salmonicida. Syst Appl Microbiol 22:403-411 [ Links ]
23.Huss VAR, Festl H, Schleifer KH (1983) Studies on the spectrophotometric determination of DNA hybridization from renaturation rates. Syst Appl Microbiol 4:184-192 [ Links ]
24. Huys G, Coopman R, Janssen P, Kersters K (1996) High-resolution genotypic analysis of the genus Aeromonas by AFLP fingerprinting. Int J Syst Bacteriol 46:572-580 [ Links ]
25. Huys G, Kämpfer P, Altwegg M, Kersters I, Lamb A, Coopman R, Lüthy-Hottenstein J, Vancanneyt M, Janssen P, Kersters K (1997) Aeromonas popoffii sp. nov., a mesophilic bacterium isolated from drinking water production plants and reservoirs. Int J Syst Bacteriol 47:1165-1171 [ Links ]
26. Huys G, Kämpfer P, Swings J (2001) New DNA-DNA hybridization and phenotypic data on the species Aeromonas ichthiosmia and Aeromonas allosaccharophila: A. ichthiosmia Schubert et al. 1990 is a later synonym of A. veronii Hickman-Brenner et al. 1987. Syst Appl Microbiol 24:177-182 [ Links ]
27. Jahnke K.-D (1992) Basic computer program for evaluation of spectroscopic DNA renaturation data from GILFORD System 2600 spectrometer on a PC/XT/AT type personal computer. J Microbiol Methods 15:61-73 [ Links ]
28. Janda JM (2001) Aeromonas and Plesiomonas. In: Sussman M (ed) Molecular medical microbiology. Academic Press, San Diego, USA, pp 1237-1270 [ Links ]
29. Janda JM, Abbott SL, Khashe S, Kellogg GH, Shimada T (1996) Further studies on biochemical characteristics and serologic properties of the genus Aeromonas. J Clin Microbiol 34:1930-1933 [ Links ]
30. Joseph SW, Carnahan A (1994) The isolation, identification, and systematics of the motile Aeromonas species. Annu Rev Fish Dis 4:315-343 [ Links ]
31. Kaznowski A (1998) Identification of Aeromonas strains of different origin to the genomic species level. J Appl Microbiol 84:423-430 [ Links ]
32. Kimura T (1969) A new subspecies of Aeromonas salmonicida as an etiological agent of furunculosis on "sakuramasu" (Oncorhynchus masou) and pink salmon (O. gorbuscha) rearing for maturity. Part 1. On the morphological and physiological properties. Fish Pathol 3:34-44 [ Links ]
33. Kimura T (1969) A new subspecies of Aeromonas salmonicida as an etiological agent of furunculosis on "sakuramasu" (Oncorhynchus masou) and pink salmon (O. gorbuscha) rearing for maturity. Part 2. On the serological properties. Fish Pathology 3:45-52 [ Links ]
34. Kozinska A, Figueras MJ, Chacón MR, Soler L (2002) Phenotypic characteristics and pathogenicity of Aeromonas genomospecies isolated from common carp (Cyprinus carpio L.). J Appl Microbiol 93:1034-1041 [ Links ]
35. Lund V, Jenssen LM, Wesmajervi MS (2002) Assessment of genetic variability and relatedness among atypical Aeromonas salmonicida from marine fishes, using AFLP-fingerprinting. Dis Aquat Organ 50:119-126 [ Links ]
36. Martínez-Murcia AJ (1999) Phylogenetic positions of Aeromonas encheleia, Aeromonas popoffii, Aeromonas DNA hybridization group 11 and Aeromonas group 501. Int J Syst Bacteriol 49:1403-1408 [ Links ]
37. Martínez-Murcia AJ, Antón AI, Rodriguez-Valera F (1999) Patterns of sequence variation in two regions of the 16S rRNA multigene family of Escherichia coli. Int J Syst Bacteriol 49:601-610 [ Links ]
38. Martínez-Murcia AJ, Benlloch S, Collins MD (1992) Phylogenetic interrelationships of members of the genera Aeromonas and Plesiomonas as determined by 16S ribosomal DNA sequencing: lack of congruence with results of DNA-DNA hybridizations. Int J Syst Microbiol 42:412-421 [ Links ]
39. Martínez-Murcia AJ, Borrell N, Figueras MJ (2000) Typing of clinical and environmental Aeromonas veronii strains based on the 16S-23S rDNA spacers. FEMS Immunol Med Microbiol 28:225-232 [ Links ]
40. Millership SE (1996) Identification. In: Austin B, Altwegg M, Gosling PJ, Joseph SW (ed) The genus Aeromonas. John Wiley and Sons, New York, pp 85-108 [ Links ]
41. Noterdaeme L, Bigawa S, Steigerwalt AG, Brenner DJ, Ollevier F (1996) Numerical taxonomy and biochemical identification of fish associated motile Aeromonas spp. Syst Appl Microbiol 19:624-633 [ Links ]
42. Oakey HJ, Ellis JT, Gibson LF (1996) A biochemical protocol for the differentiation of current genomospecies of Aeromonas. Zentralbl Bakteriol 284:32-46 [ Links ]
43. Pavan ME, Abbott SL, Zorzópulos J, Janda JM (2000) Aeromonas salmonicida subsp. pectinolytica subsp. nov., a new pectinase-positive subspecies isolated from a heavily polluted river. Int J Syst Evol Microbiol 50:1119-1124 [ Links ]
44. Popoff M (1984) Genus III. Aeromonas Kluyver and Van Neil 1936, 398AL. In: Krieg NR, Holt JG (ed) Bergey"s manual of systematic bacteriology. Baltimore, Williams & Wilkins, pp 545-548 [ Links ]
45. Popoff M, Coynault C, Kiredjan M, Lemelin M (1981) Polynucleotide sequence relatedness among motile Aeromonas species. Curr Microbiol 5:109-114 [ Links ]
46. Saavedra MJ, Guedes-Novais S, Alves A, Rema P, Tacão M, Correia A, Martínez-Murcia A (2004) Resistance to b-lactam antibiotics in Aeromonas hydrophila isolated from rainbow trout (Oncorhynchus mykiss). Int Microbiol 7:207-211 [ Links ]
47. Smith IW (1963) The classification of "Bacterium salmonicida". J Gen Microbiol 33:263-274 [ Links ]
48. Soler L, Figueras MJ, Chacón MR, Guarro J, Martinez-Murcia AJ (2003) Comparison of three molecular methods for typing Aeromonas popoffii isolates. Ant Leeuw 83:341-349 [ Links ]
49. Soler L, Yañez MA, Chacon MR, Aguilera-Arreola MG, Catalán V, Figueras MJ, Martínez-Murcia AJ (2004) Phylogenetic analysis of the genus Aeromonas based on two housekeeping genes. Int J Syst Evol Microbiol 54:1511-1519 [ Links ]
50. Sørum H, Holstad G, Lunder T, Hastein T (2000) Grouping by plasmid profiles of atypical Aeromonas salmonicida isolated from fish, with special reference to salmonid fish. Dis Aquat Organ 41:159-171 [ Links ]
51. Stackebrandt E, Frederiksen W, Garrity GM, Grimont PAD, Kämpfer P, Maiden MCS, Nesme X, Rosselló-Mora R, Swings J, Trüper HG, Vauterin L, Ward AC, Whitman WB (2002) Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int J Syst Evol Microbiol 52:1043-1047 [ Links ]
52. Wiklund T, Dalsgaard I (1998) Occurrence and significance of atypical Aeromonas salmonicida in non-salmonid and salmonid fish species: a review. Dis Aquat Organ 32:49-69 [ Links ]
53. Yamamoto S, Kasai H, Arnold DL, Jackson RW, Vivian A, Harayama S (2000) Phylogeny of the genus Pseudomonas: intrageneric structure reconstructed from the nucleotide sequences of gyrB and rpoD genes. Microbiology 146:2385-2394 [ Links ]
54. Yañez MA, Catalán V, Apráiz D, Figueras MJ, Martínez-Murcia AJ (2003) Phylogenetic analysis of members of the genus Aeromonas based on gyrB gene sequences. Int J Syst Evol Microbiol 53:875-883 [ Links ]

