










 Custom services
Custom services
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
La evidencia acumulada durante los últimos años ha establecido que la vía Wnt/β-catenina es crucial para la formación ósea y el mantenimiento de la homeostasis esquelética1,2. Las proteínas Wnt ejercen sus funciones celulares activando diferentes rutas de señalización, comúnmente denominadas vía canónica y vías no canónicas3. La primera actúa controlando la cantidad de β-catenina no asociada a cadherina, mientras que las otras rutas no requieren la presencia de β-catenina4. En la actualidad, la vía de señalización mediada por la β-catenina es la mejor estudiada y comprendida. La activación de la ruta Wnt/?catenina comienza en la membrana celular con la unión de ciertos ligandos Wnt, tales como Wnt3a, a los receptores transmembrana de la familia Frizzled. Esta unión recluta el correceptor LRP5/6 (low-density-lipoprotein receptor-related protein 5/6), para formar un complejo ternario que desestabiliza un conglomerado citoplásmico de proteínas que, de lo contrario, fosforilarían la β-catenina del citoplasma para su destrucción en el proteasoma5-7. De modo que, tras la unión del ligando al receptor, la β-catenina no es fosforilada ni destruida, y, por lo tanto, puede acumularse en el citoplasma, desde donde se trasladará al núcleo. Allí se une al factor de trascripción TCF/LEF (T-cell factor/lymphoid enhancer factor) e induce la expresión de genes diana8.
La demostración inicial de la implicación de la vía canónica de Wnt en la osteogénesis fue proporcionada por la identificación de las mutaciones de pérdida de función (LOF) y ganancia de función (GOF) del correceptor LRP5, como responsables del síndrome de osteoporosis-pseudoglioma y del fenotipo hereditario de masa ósea elevada, respectivamente9,10. Los fenotipos óseos de estas mutaciones se pudieron reproducir en modelos de ratón con la función del receptor LRP5/6 genéticamente modificada. Estos estudios pusieron de manifiesto el papel osteo-anabolizante de la señalización de Wnt/β-catenina11. Más recientemente, estudios realizados con ratones en los que se había manipulado la actividad de la β-catenina en células de linaje osteoblástico han establecido que la β-catenina aumenta la masa ósea a través de diferentes mecanismos dependiendo de la etapa de diferenciación en la que se encuentre la célula osteoblástica12-14. De forma que la GOF de la β-catenina exclusivamente en las células precursoras estimula su proliferación y maduración, pero suprime el destino alternativo hacia la diferenciación condrocítica, produciendo así un aumento de la masa ósea15. Por otro lado, cuando la GOF de β-catenina se produce en una etapa posterior de la diferenciación, se logra también una masa ósea elevada, pero esta ocurre a espensas de un mecanismo indirecto, esto es, la β-catenina induce la expresión de osteoprotegerina (OPG) en el osteoblasto, y la OPG atenúa la osteoclastogénesis16,17.
La participación indirecta de los osteoclastos como mediadores de algunos de los efectos de la señalización Wnt/β-catenina en las células de estirpe osteoblástica, planteó la cuestión de si esta vía podría tener tambien un papel directo en la función de los osteoclastos cuando se activa en ellos. Otero et al.18 y Albers et al.19 eliminaron β-catenina de los precursores de osteoclastos utilizando el modelo de ratón LisozimaMCre (LysMCre), y encontraron una disminución de la masa ósea en el compartimiento trabecular, con un aumento paralelo en el número de osteoclastos, mostrando así que la β-catenina de los osteoclastos está, de hecho, implicada en la homeostasis ósea. Estos autores, atribuyeron el aumento del número de osteoclastos a un incremento exclusivo en la osteoclastogénesis, sin considerar posibles efectos sobre la apoptosis. Creemos que es importante aclarar esta cuestión, ya que la β-catenina es parte de una vía de señalización que actualmente es considerada como un objetivo terapéutico interesante. Para obtener una idea de este problema, hemos generado un modelo de ratón similar, en el que hemos analizado su fenotipo, y tambien posibles efectos sobre la apoptosis de los osteoclastos.
MATERIAL Y MÉTODOS
1. Reactivos
La proteina WNT3a, la proteina Dickkopf 1 (DKK1), el factor estimulante de colonias de macrófagos (M-CSF) y el activador de receptor soluble de proteínas recombinantes del ligando kappa-B de factor nuclear (sRANKL) se adquirieron de R&D Systems (Minneapolis, EE.UU.).
2. Ratones mutantes
Todos los procedimientos con animales se llevaron a cabo de acuerdo con las normas de la Unión Europea y los principios 3R. Los experimentos fueron revisados y aprobados por el Comité de Bioética de la Universidad de Cantabria. El ratón con el alelo de β-catenina flaqueado por la secuencia flox20 y la línea LysMCre21 han sido descritos previamente y se obtuvieron del Laboratorio Jackson. Los ratones se genotiparon mediante PCR, utilizando el ADN genómico extraído de biopsia de la cola. Los cebadores utilizados para la detección de LysMCre fueron los siguientes; foward GCGGTCTGGCAGTAAAAACTATC y reverse: GTGAAACAGCATTGCTGTCACTT, y el tamaño del producto 102 pb. Reverse: CACCATGTCCTCTGTCTATTC, y el tamaño del producto en este caso sería de 324 pb. Los ratones experimentales se generaron mediante una estrategia de apareamiento consistente en dos pasos. Los ratones heterocigotos LysMCre fueron cruzados con ratones cuyo gen de la β-catenina estaba flanqueado por secuencias flox en homocigosis (β-cateninf/f). Para generar ratones homocigóticos para el alelo condicional de β-catenina con y sin el alelo Cre, los ratones β-cateninf / +; LysMCre se cruzaron con los ratones β-cateninf /f.
3. Micro-TAC
Se realizó un análisis de micro-TAC de la quinta vértebra lumbar después de disecar los huesos, limpiarlos, fijarlos en etanol, cargarlos en tubos de exploración de 12,3 mm de diámetro y obtener una imagen (μCT40, Scanco Medical. Basserdorf, Suiza). Los escaneos se integraron en imágenes voxel 3-D (matrices de 1.024 x 1.024 píxeles para cada pila planar individual) y se usó un filtro gaussiano (sigma=0,8, soporte=1) para reducir el ruido de la señal. Se aplicó un umbral de 200 a todos los escaneos analizados. Los escaneos se realizaron a una resolución media (E=55 kVp, I=145 μA, tiempo de integración=200 ms). Se escaneó todo el cuerpo vertebral con una orientación transversal que excluía los pedículos y los procesos articulares. El análisis manual excluyó el hueso cortical del análisis. Todas las mediciones trabeculares se realizaron dibujando manualmente contornos cada 10 a 20 cortes y utilizando el conteo de voxel para el volumen óseo por volumen de tejido y los índices de transformación de distancia de relleno esférico sin suposiciones sobre la forma del hueso como varilla o placa para microarquitectura trabecular. El espesor cortical se midió en la mitad de la diáfisis femoral.
4. Histología ósea
Las vértebras lumbares (L1, L2 y L3) y el fémur izquierdo se fijaron en paraformaldehído al 4% durante toda la noche a 4ºC y se descalcificaron durante ocho a nueve días en EDTA al 9% (pH 7,4) antes de la inclusión en parafina. El examen histomorfométrico se realizó en secciones longitudinales de 7 μm del fémur para hueso cortical y en las secciones frontales de las vértebras para hueso esponjoso.
5. Cultivos celulares
Para cuantificar las células progenitoras de los osteoclastos, la médula ósea (MO) se purgó de los huesos largos y se sembró a una densidad de 50.000 células/cm2 en placas de 48 pocillos. Tras permanecer 4-5 días en la placa de cultivo con medio α-MEM (Invitrogen), suplementado con 10% de SFB, 1% de PSG, 30 ng/ml de M-CSF y 30 ng/ml de sRANKL (R & D Systems), los osteoclastos se fijaron con 10% de formalina durante 15 min y teñida para fosfatasa ácida resistente a tartrato (TRAP). Los osteoclastos se cuantificaron teniendo en cuenta las células multinucleares y positivas para la tinción de TRAP. Para el resto de los cultivos, se usaron células de médula ósea purificadas, es decir no adheridas a la placa. Los macrófagos y osteoclastos se desarrollaron a partir de células de la médula ósea (MO) no adheridas a la placa de cultivo, y cultivadas durante 4 días en presencia de M-CSF (130 ng/ml) para obtener macrófagos, o durante 4 días en presencia de M-CSF (30 ng/ml) y 30 ng/ml de RANKL para obtener osteoclastos.
6. PCR cuantitativa en tiempo real (Quantitative real-time PCR, qRT-PCR)
El ARN total se extrajo con reactivos TRIzol (Life Tecnologies). Se usó 1 μg de ARN total para producir ADNc de primera cadena usando la enzima m-MLV RT (Invitrogen). La qRT-PCR se llevó a cabo utilizando PreMix Ex taq (Takara) y los datos se analizaron utilizando el software Biorad. Los cebadores y las sondas para β-catenina [Mm01350385_g1 (fam)] y gapdh [Mm99999915_g1 (vic)] fueron fabricados por el servicio TaqMan Gene Expression Assays (Applied Biosystems). Los niveles relativos de expresión de ARNm se normalizaron con el gen de la proteína ribosómica S2 mediante el método ΔCt22.
7. Analisis por Western blot
Los niveles de proteína de β-catenina y β-actina se analizaron usando un anticuerpo monoclonal de ratón que reconoce β-catenina (BD Biosciences), y un anticuerpo monoclonal de ratón que reconoce β-actina (Sigma-Aldrich).
8. Analisis de apoptosis
Se obtuvieron osteoclastos derivados de células de la MO no adheridas a la placa de cultivo extraídos de los ratones Catnbf/f;LysMCre y sus controles. Una vez desarrollados los osteoclastos en la placa de cultivo se trataron con WNT3a. Tras 24 horas de tratamiento, los cultivos se fijaron y se sometieron a tinción de TUNEL y TRAP. Se cuantificaron el número total de osteoclastos y el número de osteoclastos apoptóticos en cada placa. Se consideraron apoptóticos cuando al menos uno de sus núcleos era TUNEL positivo. El método TUNEL se realizó utilizando el kit de detección de fragmentación de ADN FragEL (EMD Chemicals. San Diego, California, EE.UU.) antes de la tinción para TRAP. Se enumeraron las células multinucleares TRAP positivas y TUNEL positivas.
La actividad de la caspasa-3 se midió determinando la degradación del sustrato fluorométrico DEVD (Biomol Research Laboratories, Plymouth Meeting. Pensilvania, EE.UU.), y se midió la concentración de proteína usando un kit compatible con detergente Bio-Rad (Bio-Rad Laboratories, Hercules. California, EE.UU.).
RESULTADOS
1. La eliminación específica de la β-catenina de los precursores de los osteoclastos
La inactivación condicional de la β-catenina (Catnb) en células de linaje osteoclástico se realizó cruzando ratones que albergaban un alelo de β-catenina floxeado (Catnbf)20 con ratones que expresaban la enzima Cre recombinasa bajo el control de los elementos reguladores del gen LisozimaM (LysMCre)21. Esta línea Cre induce la recombinación del alelo floxeado específicamente en las células del linaje monocito-macrófago y neutrófilos. A partir de este cruce, se generaron dos cohortes (machos y hembras) de animales; una cohorte de experimentación, con genotipo Catnbf/f;LysMCre y otra control con genotipo Catnbf/f.
Los ratones Catnbf/f;LysMCre nacieron en la proporción mendeliana esperada, con peso corporal similar (Figura 1A) a los controles de la misma camada y no mostraron ningún fenotipo evidente. La escisión del gen de β-catenina se confirmó por qRT-PCR. Los niveles de ARNm de β-catenina se analizaron en cultivos ex vivo de macrófagos y osteoclastos derivados de médula ósea (MO). Los macrófagos y osteoclastos obtenidos de ratones Catnbf/f;LysMCre exhibieron una disminución del 70% y del 60% en la expresión de β-catenina, respectivamente (Figura 1B). Atribuimos la eficacia limitada de la recombinación a la presencia de células de linajes distintos al mieloide en la placa de cultivo que expresaban niveles normales de β-catenina. Como se esperaba, el nivel de expresión de ARNm de β-catenina en riñón, hígado y bazo fue indistinguible entre los dos genotipos (Figura 1C).

Figura 1 Los ratones Catnbf/f;LysM-cre+/- expresan niveles más bajos de β-catenina en macrófagos y osteoclastos. A: peso corporal total de dos cohortes (15 animales por grupo) de ratones Catnbf/f;LysM-cre+/-y su camada control Catnbf/f de 28 semanas de edad. B: análisis cuantitativo de ARNm por PCR en tiempo real (Real Time-PCR) en macrófagos y osteoclastos desarrollados a partir de células de MO no adherentes cultivadas en presencia de M-CSF durante 4 días, y M-CSF más RANKL durante 5 días, respectivamente. C: análisis cuantitativo de ARNm de los tejidos blandos (indicados) obtenidos de ratones de 28 semanas de edad (n=5)
2. Los animales carentes de β-catenina en los precurores de los osteoclastos presentan menor grosor del hueso cortical
A las 28 semanas de vida se sacrificó una cohorte de 15 animales por sexo y genotipo y se examinó la arquitectura ósea de la quinta vértebra lumbar y el fémur derecho mediante micro-TC. El análisis reveló un grosor reducido del hueso cortical (Figura 2A), tanto en los machos como en las hembras Catnbf/f;LysMCre. Sin embargo, la ausencia de β-catenina en los precursores de los osteoclastos no alteró significativamente la masa ósea trabecular, ni en las vértebras, ni en el fémur (Figura 2B). Tampoco se vio afectado el espacio intertrabecular, el grosor trabecular y el número de trabeculas (no mostrado). Estos resultados sugieren que la señalización de Wnt/β-catenina en los osteoclastos es importante para el mantenimiento de la masa ósea cortical.

Barras: valores expresados como media ± desviación estándar; Oc: osteoclasto; *p=0,05.
Figura 2 Los ratones Catnbf/f;LysM-cre+/-tienen menor masa ósea cortical. Mediciones de tomografía micro computarizada (μ-CT) realizadas en huesos de ratones de 28 meses (n=12-15 ratones por grupo). A: espesor cortical (Cortical th) determinado en fémures. B: BV/TV, volumen óseo por volumen total de tejido determinado en L5 y fémur derecho (solo se muestran las hembras)
3. Los ratones Catnbf/f;LysMCre presentan más osteoclastos en el endostio y más progenitores osteoclásticos en la médula ósea (MO)
A continuación, quisimos cuantificar el número de osteoclastos presentes en la superficie del hueso cortical. Lo que encontramos fue que, de acuerdo con un espesor cortical disminuido, el número de osteoclastos en la superficie endocortical del fémur de los ratones Catnbf/f;LysMCre estaba aumentado, en comparación con los animales control (Figura 3A). El hueso trabecular vertebral parecía mostrar la misma tendencia. Sin embargo, la diferencia no fue significativa (p=0,06).

Barras: valores expresados como media ± desviación estándar; Oc: osteoclasto; *p=0,05.
Figura 3 Los ratones Catnbf/f; LysM-cre+/-tienen mas osteoclastos que la camada control. A: análisis histomorfométrico de secciones longitudinales descalcificadas de fémur y vértebras L1-L3 de ratones hembra de 28 meses (n=5 ratones por grupo). Las fotomicrografías (x40) muestran áreas representativas de la superficie ósea endosteal tras la tinción de TRAP. B: número de células positivas para TRAP desarrolladas a partir de células de MO, obtenidas de fémures de ratones de 28 semanas de edad, y cultivadas en presencia de M-CSF y RANKL durante 5 días. Se realizaron cultivos por triplicado de cada uno de tres animales por separado. Cada barra representa triplicados de cada ratón, n=3 por grupo. Las fotomicrografías (x40) muestran áreas representativas de la placa de cultivo
Para examinar si el mayor número de osteoclastos en el hueso podría explicarse por un aumento de la osteoclastogénesis, se cuantificó el número de estos progenitores en la MO. Con este propósito, las células de la MO obtenidas de los huesos largos de ratones de 28 semanas de edad se cultivaron en presencia de RANKL y M-CSF durante 5 días. Se utilizaron tres ratones por genotipo y se realizaron determinaciones por triplicado para cada uno de ellos. El número de osteoclastos que se desarrolló en los cultivos procedentes de ratones Catnbf/f;LysMCre mostraron una morfología normal (Figura 3B). Sin embargo, la cantidad fue dos veces mayor que en los cultivos procedentes de la camada control (426±18 por pocillo frente a 238±77, p=0,015) (Figura 3B). Este resultado sugiere que la señalización de Wnt/β-catenina en los precursores de osteoclastos y su descendencia atenúa la cantidad de osteoclastos maduros.
4. El efecto proapoptótico de WNT3a requiere la presencia de β-catenina
Para examinar los mecanismos celulares a través de los cuales Wnt/β-catenina disminuía el número de osteoclastos, primero determinamos si la adición de WNT3a a la placa de cultivo de los precursores de osteoclastos interfería con el desarrollo de los mismos. Como se muestra en la figura 4A, la presencia de WNT3a en el medio de cultivo disminuyó el número de osteoclastos inducidos por la presencia de M-CSF y RANKL. Además, este hecho parecía ser dosis dependiente. El análisis por Western blot confirmó un aumento de los niveles de β-catenina en los osteoclastos expuestos a WNT3a (Figura 4B). Además, el tratamiento del cultivo con DKK1, un inhibidor del correceptor LRP5/6 evitó el aumento de β-catenina inducido por WNT3a. El conjunto de estos hallazgos indica que WNT3a inhibe el desarrollo de los osteoclastos mediante la estimulación de la vía canónica de Wnt.
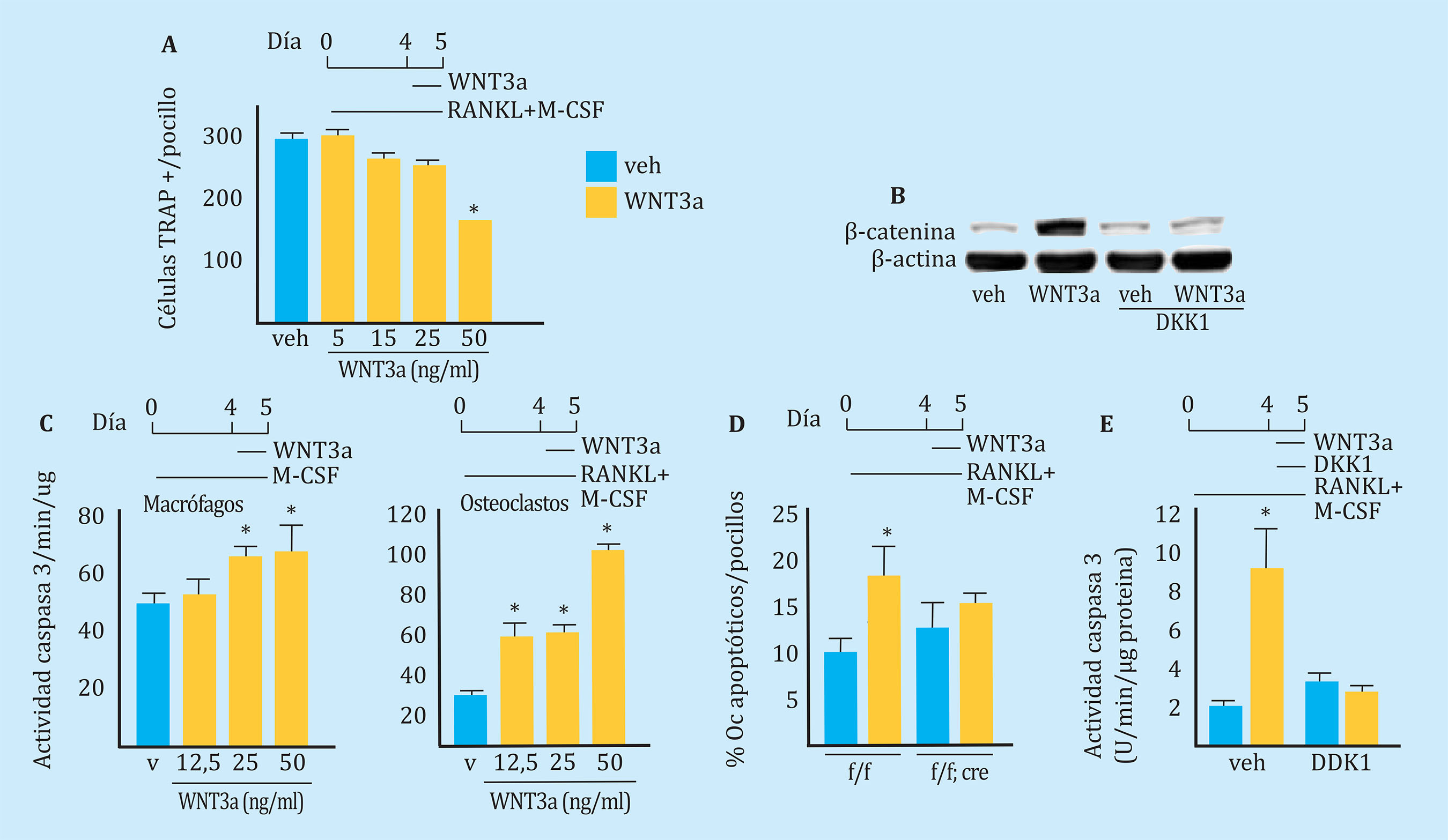
Barras: valores expresados como media ± desviación estándar; Oc: osteoclasto; * p=0,05.
Figura 4 WNT3a induce la apoptosis de los osteoclastos a través de la vía canónica de Wnt. A: número de células positivas para TRAP generadas a partir de células de MO no adheridas a la placa obtenidas de ratones C57BL/6 y cultivados con M-CSF, RANKL y placebo (veh) o dosis crecientes de proteína recombinante WNT3a como se indica. B: análisis de Western blot de β-catenina en osteoclastos maduros tratados con veh, WNT3a, DKK1 o ambos. C: actividad de caspasa 3 en macrófagos y osteoclastos maduros generados a partir de células de MO no adherentes obtenidas de ratones C57BL/6 y tratados con veh o con diferentes dosis de WNT3a durante 16 horas. D: ensayo TUNEL realizado en cultivos de osteoclastos maduros generados a partir de células de MO de ratones Catnbf/f;LysM-cre cultivadas con M-CSF y RANKL durante 5 días y tratados con veh o WNT3a (50 ng/ml) durante 24 horas. E: actividad de la caspasa 3 en osteoclastos maduros generados a partir de células de MO no adherentes de ratones C57BL/6, y tratada con veh, WNT3a (50 ng/ml) o DKK1 (1 μg/ml) durante 24 h. AFU, unidades fluorescentes arbitrarias
Es bien sabido que la vida media de los osteoclastos es muy breve, y que mueren por apoptosis. A continuación examinamos el efecto de la señalización de Wnt en la apoptosis de los macrófagos y de los osteoclastos.
Como se muestra en la figura 4C, se determinó la apoptosis midiendo la actividad de la caspasa-3 tras la administración de dosis crecientes de WNT3A a los cultivos de macrófagos y de osteoclastos. Los resultados mostraron que WNT3a inducía la apoptosis tanto en los macrófagos, como en los osteoclastos. El efecto proapoptótico de WNT3 era también dependiente de la dosis utilizada. El efecto de WNT3a sobre la apoptosis de los osteoclastos también se analizó mediante TUNEL (Figura 4D), y, de la misma forma que se observó en el experimento anterior, vimos que el tratamiento con WNT3a aumentaba el porcentaje de osteoclastos TUNEL positivos. Sin embargo, la presencia de WNT3 no tuvo ningun efecto deletéreo sobre los cultivos de osteoclastos obtenidos de los ratones Catnbf/f;LysMCre (Figura 4D), carentes de β-catenina. Igualmente, la adición de DKK1 a los cultivos anuló las acciones proapoptóticas de WNT3a (Figura 4D), indicando que el efecto proapoptótico de WNT3a requiere la presencia de β-catenina. En conjunto, estos resultados apoyan la hipótesis de que la vía de señalización canónica de Wnt/β-catenina ejerce efectos proapoptóticos en los osteoclastos.
DISCUSIÓN
En este trabajo hemos analizado las características óseas de animales que carecen de β-catenina en las células del linaje monocito/macrófago, que son los precursores de los osteoclastos. Estos animales muestran un grosor cortical reducido que se asocia a mayor cantidad de osteoclastos en la superficie del endostio y un mayor número de progenitores osteoclásticos en la MO. Además, demostramos que la estimulación de la señalización de Wnt/β-catenina en los osteoclastos atenúa la cantidad de osteoclastos desarrollados inducida por la presencia de M-SCF y RANKL, y promueve su apoptosis.
Otero et al.18 y Albers et al.19 utilizaron la misma línea LysMCre para eliminar la β-catenina de los precursores de osteoclastos. Ambos grupos describieron una disminución de la masa ósea en el compartimiento trabecular, con un aumento paralelo en el número de osteoclastos, que atribuyeron a un aumento exclusivo de la osteoclastogénesis. Sin embargo, no abordaron la posibilidad de que una disminución en la apoptosis osteoclástica estuviera también implicada. En nuestro estudio, demostramos un aumento de los osteoclastos en la superficie endocortical del hueso femoral, y un aumento en el número de osteoclastos generados en los cultivos celulares obtenidos de animales Catnbf/f;LysMCre, en comparación con los controles. También observamos que la estimulación de la señalización de Wnt/β-catenina en las células de MO obtenidas de ratones salvajes disminuye el número de osteoclastos desarrollados en la placa de cultivo. Nuestros resultados, por lo tanto, coinciden con los de Otero et al.18 y Albers et al.19. Además, nosotros hemos abordado la cuestión de una posible implicación de la apoptosis osteoclástica como parte del mecanismo de acción subyacente a los hallazgos fenotípicos observados. Nuestros hallazgos, en este sentido, indican que la disminución en el número de osteoclastos inducida por la activación de Wnt/β-catenina se debe claramente a la estimulación de la apoptosis de los macrófagos y osteoclastos.
Varios estudios han demostrado que alteraciones en la supervivencia de los osteoclastos modifican la masa ósea23-26. De hecho, está bien establecido que los estrógenos protegen el esqueleto, en parte, a través de los efectos proapoptóticos sobre los osteoclastos27,28. La eliminación del receptor de estrógeno alfa en las células del linaje osteoclástico, similar a la eliminación de β-catenina, aumenta el número de osteoclastos y disminuye la masa ósea. A diferencia de los estrógenos, los glucocorticoides promueven la pérdida de masa ósea, al menos en parte, a través de la prolongación de la vida útil de los osteoclastos29.
Las proteínas Wnts ejercen un efecto positivo sobre la supervivencia de los osteoblastos y también sobre los progenitores osteoblásticos que aún no se han comprometido30. Esta acción antiapoptótica de las proteínas Wnts sobre los osteoblastos se ha postulado como uno de los mecanismos por los cuales la señalización de Wnt aumenta la masa ósea31. Aunque la vía Wnt/β-catenina es mejor conocida por sus efectos pro-supervivencia, también puede ejercer acciones proapoptóticas. Por ejemplo, la apoptosis de los cardiomioblastos de rata inducida por la reoxigenación tras la hipoxia está regulada por WNT3a, a través de un mecanismo dependiente de β-catenina32. Además, en línea con estos hallazgos, la señalización de Wnt/β-catenina disminuye la invasividad celular del melanoma33, potenciando la expresión de proteínas proapoptóticas, como BIM y PUMA, y disminuyendo los niveles de proteínas antiapoptóticas, como MCL34.
En nuestro estudio, mediante el análisis de actividad de la caspasa, o ensayos de TUNEL, hemos encontrado que WNT3a induce la apoptosis de los osteoclastos. Este efecto es contrario a su efecto de pro-supervivencia previamente mencionado sobre los osteoblastos30. Curiosamente, al igual que Wnts, los estrógenos y los glucocorticoides ejercen efectos opuestos sobre la apoptosis de osteoblastos y osteoclastos29,35,36.
Para concluir, nuestros hallazgos sugieren que los efectos inhibidores de la β-catenina sobre los osteoclastos se deben a mecanismos proapoptóticos y apoyan la hipótesis de que los efectos osteoprotectores de la vía canónica de Wnt son también el resultado de la acción directa a través de las células del linaje osteoclástico. Por lo tanto, la señalización Wnt/β-catenina en el entorno óseo tiene un efecto osteoprotector ejercido tanto a través de las células de linajes osteoblásticos como osteoclásticos.

