










 Custom services
Custom services
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El oxígeno es necesario para la producción de energía celular y está implicado en numerosos procesos, como la activación enzimática, la señalización molecular y la regulación de la expresión de genes1. También en la angiogénesis, el mantenimiento de las células madre hematopoyéticas y la formación ósea2. De hecho, cambios en la presión parcial de oxígeno pueden influir en la función de los osteoblastos y osteoclastos3. En hipoxia desciende la formación y la mineralización del hueso, mientras que aumenta la resorción4-6. En sentido contrario, la hiperoxia podría tener un efecto beneficioso para el hueso. El tratamiento con oxígeno a alta concentración en cámara hiperbárica (CH) ha demostrado su utilidad en la osteomielitis y la osteonecrosis de mandíbula causada por radioterapia o por el uso de bisfosfonatos7-9. La CH acelera la diferenciación osteogénica de células mesenquimales y disminuye la activación de osteoclastos10-12.
En este trabajo hemos querido analizar las acciones del oxígeno a alta concentración en CH sobre la expresión de genes relacionados con el metabolismo óseo en líneas celulares osteoblásticas y hueso humano5,6,13,14.
Material y métodos:
Líneas celulares
Se utilizaron dos líneas celulares osteoblásticas Saos-2 y Super-Saos. Saos-2, derivada de un osteosarcoma humano. Super-Saos es una línea generada en nuestro laboratorio, derivada de la anterior y con alta capacidad de expresar el gen de la esclerostina (SOST)15. Ambas líneas fueron cultivadas en frascos T25 con 5 ml de medio de cultivo DMEM (medio de cultivo Eagle modificado de Dulbeco) más 1% P/S (penicilina-estreptomicina) y 1% anfotericina B, y se guardaron en incubadora a 37ºC durante una semana, cambiando el medio de cultivo cada 4 días hasta cubrir entre el 60-80% de la superficie del frasco. Las placas se introdujeron en la CH (Galeazzi, Italia; 100% oxígeno; 2,4 atmósferas) durante 90 minutos por sesión recibiendo una, tres o cinco sesiones consecutivas (Figura 1). Se utilizó como grupo control la misma línea celular sometida a idénticas condiciones de cultivo, transporte y manipulación, pero sin someterse a la CH.

Fragmentos óseos
Se utilizaron fragmentos de hueso trabecular extraídos de la cabeza femoral de pacientes con cirugía de reemplazo de cadera por fractura osteoporótica. Tras la extracción, los fragmentos óseos recibieron una sola sesión de CH (Galeazzi, Italia; 100% oxígeno; 2,4 atmósferas) durante 90 minutos y posteriormente se congelaron a -70ºC. Se utilizaron como controles fragmentos de hueso sometidos a las mismas condiciones de conservación, cultivo, transporte y manipulación pero sin recibir CH. Este experimento fue aprobado por el Comité Ético de Investigación Clínica (CEIC) de Cantabria. Todos los pacientes dieron su consentimiento informado.
Extracción y cuantificación de ARN
A las 24 horas de la última sesión de CH se procedió a la extracción del ARN, tanto en las líneas celulares como en el hueso. En el proceso de homogenización en líneas celulares se realizó lavado de las muestras con tampón fosfato salino (PBS) previo al uso del TRIzol®. En el caso de los fragmentos óseos también se empleó TRIzol®, así como homogeinización durante 20-30 segundos hasta conseguir pulverizar la muestra, y posteriormente se centrifugó. En ambos casos se siguieron las recomendaciones del fabricante y se continuó con el proceso de separación, precipitación y resuspensión del ARN.
Se usó RT-PCR (reacción en cadena de la polimerasa con transcriptasa reversa) cuantitativa para detectar la expresión de los genes: SOST (gen de esclerostina), RUNX2 (proteína relacionada con el factor de transcripción 2), MMP14 (metaloproteinasa 14), HIF-1α (factor inducible por hipoxia), SIRT1 (sirtuina1), OPG (osteoprotegerina) y RANKL (ligando del activador del receptor del factor nuclear kappa-B) utilizando ensayos Taqman y siguiendo las instrucciones del fabricante. Se obtuvieron los valores del ciclo umbral (Ct) y los datos se normalizaron a la expresión de GAPDH (gliceraldehído-3-fosfato deshidrogenasa) y TBP (proteína de unión a caja TATA) utilizando el método ΔCt. Para calcular el nivel relativo de ARNm se usó la fórmula 2-ΔCt, donde ΔCt es la diferencia entre la media de los Ct de los genes normalizadores y el Ct del gen de interés.
Resultados:
Efecto de CH en la expresión de ARN en la línea celular Saos-2
No hubo diferencias en la expresión de genes en la línea celular tras una, tres o cinco sesiones de CH. Las diferencias respecto al control en 2-ΔCt tras 5 sesiones fueron de 0,71 para SOST (p=0,50), 0,89 para SIRT1 (p=0,34), 0,47 para MMP14 (p=0,18), 0,43 para HIF1α (p=0,18), 0,79 para RUNX2 (p=0,65) y 7,91 para OPG (p=0,40) (Figura 2). No se detectó expresión de RANKL.
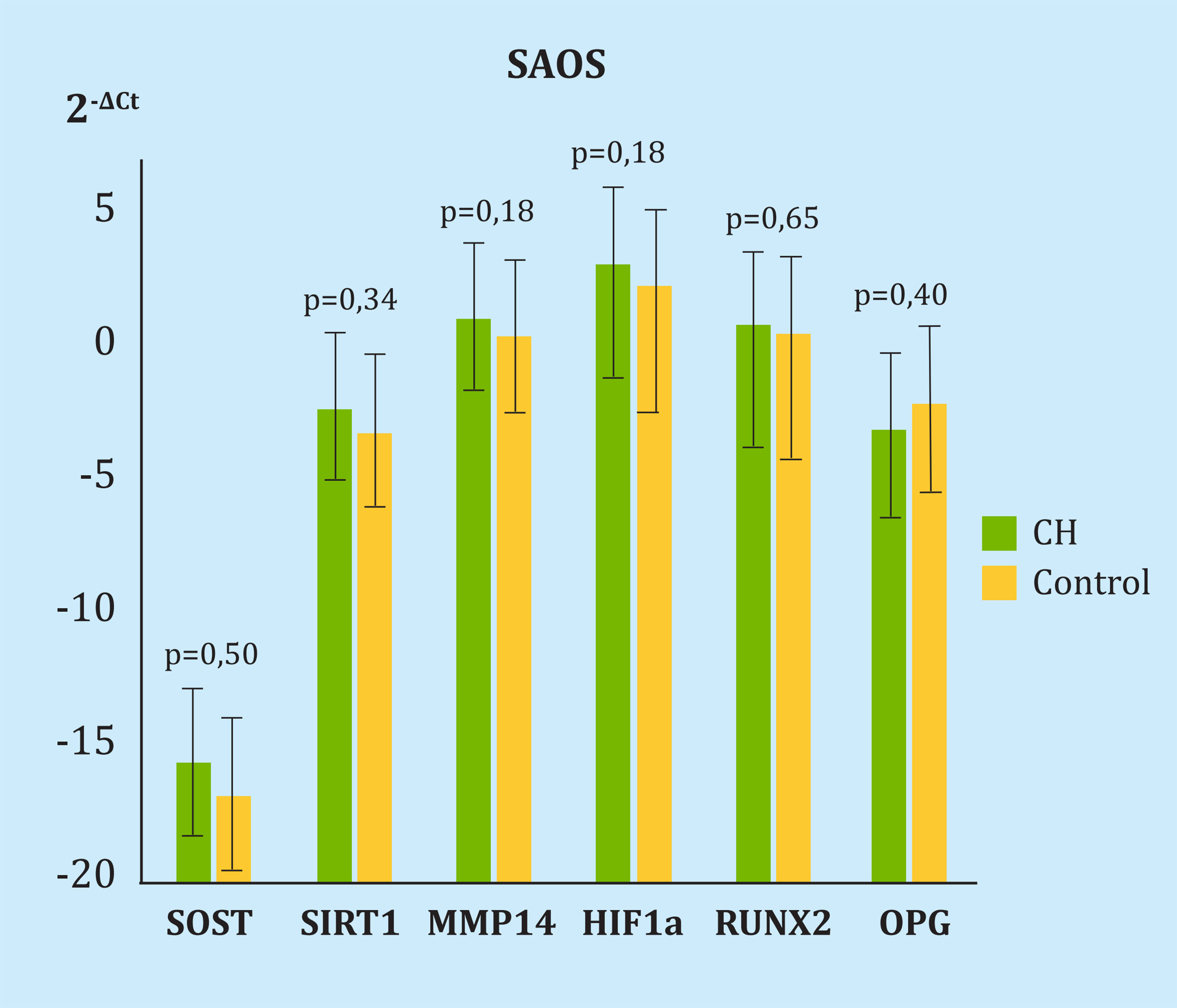
CH: grupo celular sometido a cámara hiperbárica; SOST: esclerostina; SIRT1: sirtuina1; MMP14: metaloproteinasa 14; HIF-1α: factor inducible por hipoxia 1α; RUNX2: proteína relacionada con el factor de transcripción 2; OPG: osteoprotegerina.
Figura 2. Diferencia en la expresión de genes a estudio y los housekeeping en la línea celular SAOS tras 5 sesiones de cámara hiperbárica
Efecto de CH en la expresión de ARN en la línea celular Super-Saos
Encontramos con respecto al control que la expresión de OPG disminuye 6 veces tras 5 sesiones de CH (2-ΔCt, 72 p=0,01). En el resto de genes no hubo diferencias: 2-ΔCt, de 1,03 para SOST (p=0,34), 1,46 para SIRT1 (p=0,34), 1,77 para MMP14 (p=0,18), 1,08 para HIF1α (p=0,18), 1,14 para RUNX2 (p=0,18) y 1,24 para RANKL (p=0,31) (Figura 3).

CH: grupo celular sometido a cámara hiperbárica; SOST: esclerostina; SIRT1: sirtuina1; MMP14: metaloproteinasa 14; HIF-1α: factor inducible por hipoxia 1α; RUNX2: proteína relacionada con el factor de transcripción 2; OPG: osteoprotegerina.
Figura 3. Diferencia en la expresión de genes a estudio y los housekeeping en la línea celular Super-SAOS tras 5 sesiones de cámara hiperbárica
Efecto de CH en la expresión ARN en hueso trabecular
Tampoco hubo diferencias en la expresión de los genes tras CH en el hueso, solo un modesto incremento, no significativo, en la expresión de SOST con un cambio de 2-ΔCt de 5,39 (p=0,48). En el resto de los genes las diferencias fueron de 0,92 para MMP14 (p=0,58), 1,28 para HIF1α (p=0,81), 0,72 para RUNX2 (p=0,24), 1,18 para SIRT1 (p=0,42), 1,97 para RANKL (p=0,91) y 3,9 para OPG (p=0,55) (Figura 4).

CH: grupo celular sometido a cámara hiperbárica; SOST: esclerostina; SIRT1: sirtuina1; MMP14: metaloproteinasa 14; HIF-1α: factor inducible por hipoxia 1α; RUNX2: proteína relacionada con el factor de transcripción 2; OPG: osteoprotegerina.
Figura 4. Diferencia en la expresión de genes a estudio y los housekeeping en hueso tras una única sesión de cámara hiperbárica
Conclusiones:
Se cree que la hiperoxia podría ser beneficiosa para el hueso aumentando la proliferación y diferenciación de los osteoblastos16. Al Hadi y cols.6 describieron un aumento en la expresión de colágeno tipo I y ARNm de Runx-2 en líneas celulares de osteoblastos (Saos-2) sometidas a CH durante 14 días (2,4 ATA, 97% O2, 90 min/día). La CH aumentó también la proliferación y diferenciación de osteoblastos en hueso alveolar humano17. La hiperoxia además parece disminuir la resorción ósea. El tratamiento en CH (100% O2, 2,4 ATA) redujo la expresión de RANK, NFATc1 y Dc-STAMP en el suero de pacientes y reguló también la expresión del factor inducible en hipoxia (HIF-1α)18. Otras acciones descritas del oxígeno a alta concentración (O2 100%, 2,4 ATA) son la mejora en la angiogénesis, el aumento de la vascularización en aspirado de cresta ilíaca de ratones19, mayor proliferación celular20 o aceleración en la curación de fracturas abiertas de fémur en animales de experimentación21. Sin embargo, la mayoría de estos trabajos se han realizado en modelos animales, siendo escasos los estudios en humanos. En pacientes con necrosis avascular de la cabeza femoral aumentaron los niveles séricos de OPG tras la CH (5,61±1,99 pmol/L basal, 7,90±1,9 pmol/L tras 15 sesiones, 8,97±2,07 pmol/L tras 30 sesiones; p<0,05), sin producirse cambios en los niveles de RANKL22. Tras la CH (2,5 ATA, 100% O2 durante 90 min/día) mejoró también la diferenciación osteogénica de células mesenquimales de la médula ósea en los pacientes tratados, con una regulación ascendente en Wnt3a, b-catenina y Runx2 y descendente de GSK-3b, en comparación con los aquellos que no la recibieron12. Estos mismos autores describieron también un aumento de la proteína morfogenética ósea (BMP2) y de Osterix en los enfermos tratados12.
En nuestro estudio no encontramos que el oxígeno a alta concentración en CH influya en la expresión de diferentes genes relacionados con el metabolismo óseo (SOST, SIRT1, MMP14, HIF1a, RUNX2, OPG y RNAKL). Sin embargo, queremos destacar que encontramos una ligera tendencia, no significativa, al incremento en la expresión de SOST en el hueso sometido al tratamiento. Sabemos que la tensión de oxígeno influye en la regulación de SOST y que en hipoxia (tensión de oxígeno al 1%) osteoblastos y osteocitos expresan niveles bajos de SOST y esclerostina23, quizás esto es debido a una menor expresión de prolil-hidroxilasa (PHD2) ya que se ha visto que la delección de PHD2 en osteocitos origina una menor producción de esclerostina dependiente de SIRT114. Esta vía podría ayudar a comprender mejor el mecanismo fisiopatológico a través del cual, y en sentido contrario, un entorno rico en oxígeno podría aumentar la expresión de SOST y esclerostina. De hecho, nuestro grupo ha encontrado un aumento del 25% en los niveles de esclerostina en el suero de 12 pacientes sometidos al tratamiento en CH. Sin embargo, otros trabajos son contradictorios13.
En conclusión, no parece que la hiperoxia en CH influya en la expresión de genes relacionados con el metabolismo óseo, aunque consideramos que se necesitan más estudios que profundicen en el conocimiento de las acciones del oxígeno en el hueso.

