










 Custom services
Custom services
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
En los años 60, Friedenstein describió por primera vez las células estromales de la medula ósea (Bone Marrow Stromal Cells, BMSCs) como células fibroblásticas con adherencia al plástico y capacidad de diferenciación tri-lineal, generando osteoblastos, condroblastos y adipocitos. Una fracción de estas células demostró además capacidad clonogénica cuando éstas se cultivaron en muy baja densidad (Colony Forming Units-Fibroblast, CFU-F) [1]. Más adelante se observó que esta multipotencialidad es sólo inherente a una pequeña parte de esta población celular heterogénea, correspondiente a las células madre esqueléticas (Skeletal Stem Cells, SSCs, por debajo del 0,1% de las BMSCs y alrededor del 12% de las CFU-F), que, además, son las únicas capaces de generar el estroma necesario para la generación de nichos hematopoyéticos en la médula ósea [2,3].
El tejido adiposo contiene un gran número de células adherentes, con capacidad de formar CFU-F y muchas otras características que las hacen similares a las BMSCs. En comparación con las BMSCs, este tejido es de fácil acceso, como lipoaspirados descartados en operaciones de liposucción, y el aislamiento de células estromales es relativamente sencillo [4,5]. De forma similar a las SSCs entre las BMSCs, la fracción estromal vascular del tejido adiposo (SVF) es una población heterogénea de células entre las que se encuentran las ASCs, que presumiblemente se encuentran asociadas de forma similar a las SSCs a la microvasculatura de la grasa [6], en consonancia con otros tejidos en los que las células madres adultas se encuentran asociadas a la microvasculatura en forma de pericitos [7]. Aunque diversos autores afirman que las ASCs poseen la capacidad inherente de diferenciarse a células de estirpe ósea, hasta la fecha ningún grupo de investigación ha demostrado que esto sea posible, salvo después de transdiferenciar estas células tras un tratamiento intensivo con BMPs, que ha demostrado ser un potente osteoinductor de diversos tipos celulares [8]. Pese a no haberse demostrado la utilidad clínica de las ASCs para la regeneración ósea, estas células pueden ser un modelo conveniente para estudiar la diferenciación osteoblástica in vitro, gracias a su fácil acceso y a su respuesta similar a la de las BMSCs a factores de diferenciación tri-linear in vitro [9].
En presencia de ácido ascórbico, dexametasona y β glicerol-fosfato las ASC expresan marcadores de diferenciación osteoblástica in vitro. Es el caso del sistema formado por la osteoprotegerina (OPG) y el ligando de receptor activador para el factor nuclear κB (RANKL), proteínas implicadas en el remodelado óseo. RANKL es una proteína de la familia del factor de necrosis tumoral α (TNF-α) que se expresa en la membrana de los osteoblastos, y que a su vez puede ser secretada por éstas [10]. Se une al receptor activador para el factor nuclear κB (RANK) que está presente en precursores osteoclásticos activando su diferenciación y maduración a osteoclastos [11]. OPG es un receptor señuelo secretado por los osteoblastos que se une a RANKL, impidiendo que éste se una a RANK, bloqueando por tanto la activación de los osteoclastos. Por otro lado, Runx2 es un factor de transcripción clave en la diferenciación de células osteoblásticas [12].
El factor de crecimiento del endotelio vascular o VEGF, es una molécula clave en la regulación de la proliferación de las células endoteliales. Promueve la proliferación, migración y supervivencia de estas células, así como su permeabilidad vascular [13]. La expresión de VEGF y sus receptores en BMSCs ha sido demostrada por diversos estudios en cultivos celulares [13,14]. Además, in vitro, se ha mostrado el papel del receptor VEGFR2 como clave en la diferenciación y supervivencia osteoblástica [15]. VEGF induce la diferenciación en cultivos celulares de preosteoblastos [14] y estimula su migración y proliferación [13,14]. Así, el VEGF parece estar implicado en las etapas tempranas de diferenciación ósea, tanto en la esqueletogénesis –siendo un importante factor en la formación ósea endocondral e intramembranosa– como en homeostasis adulta, promoviendo la diferenciación osteoblástica y reduciendo la diferenciación adipogénica de las BMSCs [16,17].
La proteína relacionada con la parathormona (PTH), PTHrP, es una citoquina pleiotrópica con importantes funciones en el tejido óseo [15]. Es considerada un modulador del remodelado óseo y un estimulador de la formación de hueso que promueve la diferenciación osteoblástica y su supervivencia [15], siendo por tanto un importante factor de maduración. El procesamiento postraduccional del gen de la PTHrP, genera diferentes fragmentos, entre los que destaca un fragmento N-terminal, PTHrP(1-36), que presenta gran homología con la PTH y que actúa a través del receptor 1 común para PTH/PTHrP, PTH1R [12]. La expresión de PTHrP en el linaje osteoblástico disminuye a medida que ésta se va diferenciando [20], pero el PTH1R juega un papel fundamental en los osteoblastos maduros y osteocitos, disminuyendo su apoptosis y aumentando su número en osteoblastos del periostio in vivo [18,19]. Estudios previos indican que la PTHrP podría incrementar la proliferación de osteoblastos inmaduros a través de la regulación de Ciclina D1 (promotor) y p27 (inhibidor), ambos reguladores del ciclo celular [12].
En este trabajo hipotetizamos que los efectos de VEGF y PTHrP sobre la proliferación celular de los osteoprogenitores son dependientes del estadio de diferenciación osteoblástica. Así, las poblaciones más indiferenciadas serían más sensibles a VEGF mientras que los progenitores ya comprometidos hacia la diferenciación osteoblástica responderían mejor a PTHrP(1-36). Para ello se utilizaron ASCs humanas procedentes de lipoaspirados como modelo in vitro de diferenciación osteogénica. Tras confirmar su capacidad de diferenciación in vitro por expresión génica de Runx2 y acumulación de calcio, se analizó la respuesta proliferativa a estímulos con VEGF o PTHrP(1-36) de ASCs sometidas o no a inducción osteogénica. VEGF, pero no PTHrP(1-36), indujo la capacidad proliferativa de las ASCs indiferenciadas, mientras que PTHrP (1-36), pero no VEGF, indujo la proliferación de las ASCs previamente tratadas con medio de diferenciación osteogénica, confirmando el papel diferencial de estos factores de crecimiento en distintos estadios de diferenciación.
Materiales y métodos
Aislamiento, cultivo primario y estudio funcional de ASCs
El tejido adiposo subcutáneo abdominal se obtuvo durante la cirugía de pacientes sanos mediante la técnica de liposucción. Seis mujeres se incluyeron en este estudio con una edad promedio de 40 años (rango 25-60 años). Todos los donantes dieron su consentimiento informado, de acuerdo con el protocolo clínico apropiado. Los pacientes fueron operados en el Departamento de Cirugía Plástica de HM Hospitales (Madrid, España), y la recogida de muestra de tejido fue aprobado por la Junta de Revisión Institucional/Comité Ético de Investigación Clínica de HM Hospitales (Madrid, España). El tejido adiposo fue digerido en 0,075% de solución colagenasa tipo I (Invitrogen, Life Technologies, Nueva York, EE.UU.) durante 30 minutos a 37°C, siguiendo el protocolo descrito anteriormente [4]. A continuación, las células se sembraron en superficie plástica (Corning, Nueva York, EE.UU.) durante 24 horas en Medio de Crecimiento DMAX (Invitrogen, Life Technologies, Nueva York, EE.UU.) con 10% de FBS (Sigma-Aldrich; St. Louis, Missouri, EE.UU.) y suplementado con antibióticos: penicilina (100 IU/ml) y estreptomicina (100 mg/ml) (Sigma-Aldrich; St. Louis, Missouri, EE.UU.). Las células no adherentes se retiraron, y se añadió medio fresco para el cultivo primario de la fracción de células adherentes durante 7 días. El medio de cultivo se reemplazó cada 3 días. En ese momento, las ASC fueron funcionalmente fenotipadas por citometría de flujo y se analizó su potencial de diferenciación osteogénica.
Citometría de flujo
Para el análisis de citometría de flujo, las ASC se resuspendieron en PBS (tampón fosfato salino) a una densidad de 1x106 células/ml, se fijaron con paraformaldehído (PFA) al 2% y se incubaron con anticuerpos monoclonales de ratón conjugados (FITC o PE) con anti- CD90, CD29, CD34, CD45, CD106 CD44, CD144, CD31 y KDR (BioLegend, San Diego, Caifornia, EE.UU.) durante 30 min a temperatura ambiente. Las células se lavaron 3 veces con PBS y se analizaron mediante Accuri's C6 Flow System citómetro a 488 nm y 15 mW. La dispersión frontal (FSC), dispersión lateral (SSC) y el marcador fluorescente específico (LGFL) a 488 nm y 540 nm se obtuvieron de forma automática para cada célula. Los datos se recogieron digitalmente sobre un rango dinámico de 16 millones de canales de datos digitales. La amplificación y análisis logarítmico de fluorescencia se realizó utilizando BD Accuri™ Software de Análisis C6.
Ensayos de diferenciación osteogénica
Se utilizó el medio de diferenciación osteogénico de linaje celular para evaluar el potencial de diferenciación osteogénica de ASC. Para dicha diferenciación las ASC se cultivaron durante 14 días en presencia de DMEM (Dulbecco's Modified Eagle's medium) con 10% FBS, 10-8 M de dexametasona, 0,5 mg/ml de ácido ascórbico y 0,1 M de b glicerolfosfato (Sigma-Aldrich; St. Louis, Missouri, EE.UU.). El medio se reemplazó cada 3 días, y al finalizar el periodo de 14 días se realizó la tinción histoquímica rojo de alizarina para revelar y cuantificar el número de células por campo microscópico rodeadas por matriz extracelular mineralizada teñida con rojo de alizarina. Así mismo se realizó una comprobación de la diferenciación osteoblástica mediante ensayo de actividad de la fosfatasa alcalina (ALP), que consistió en la identificación de los depósitos teñidos de rojo que indican actividad fosfatasa alcalina mediante el kit 86R de Sigma Aldrich. ASCs de seis pacientes diferentes se utilizaron en cada ensayo.
Estudios de proliferación celular
Se estudió la respuesta relacionada con la proliferación de las ASCs a factores como el VEGF (160 pM) y la PTHrP(1-36) (100 nM) en los cultivos durante 24 horas, tanto en medio de crecimiento como en medio de diferenciación osteogénica. Para la medición de la proliferación de estos ensayos se utilizó el equipo de medición xCelligent System (Roche Diagnostics, Basilea, Suiza) que, a tiempo real, midió la proliferación celular. El sistema xCelligent proporcionó medidas de proliferación a tiempo real y final a través de la lectura de unas placas de cultivo dotadas de electrodos que detectaron los cambios de morfología de las células, proporcionando un parámetro llamado Cell Index extrapolable a la proliferación celular.
Extracción de ARN total y PCR cuantitativa a tiempo real
Las ASCs, tanto en condiciones basales como las diferenciadas a osteoblastos, fueron sometidas a extracción de ARN total de un homogeneizado celular con un método estándar con tiocianato de guanidilo-fenol-cloroformo (Tri-Reagent, MCR; Cincinnati, Ohio, EE.UU.). La pureza y cuantificación del ARN total extraído se determinó mediante espectrofotometría A260/A280 (Nanodrop 2000/2000c Spectrophotometer, Thermo Scientific). La síntesis de ADNc se realizó utilizando oligonucleótidos aleatorios y una transcriptasa inversa (High capacity RNA to cDNA Kit, Applied Byosystem; Foster City, California, EE.UU.). El análisis de la expresión génica por RT-PCR a tiempo real se realizó con ADNc resultante utilizando una polimerasa activada por calor, TaqDNA (Taqman gene expression master mix, Applied Byosystem; Foster City, California, EE.UU.) y cebadores específicos humanos para RUNX2, (Hs00231692 m1) OPG (TNFRSF1 Hs 00171068), VEGF-A (Hs 00173626 m1) y VEGFR2 (Hs 00176676 m1) (Applied Byosystem; Foster City, California, EE.UU.). Tras una incubación inicial de 10 minutos a 25ºC y otra de 2 horas a 37ºC, las muestras se sometieron a ciclos a 4ºC. Los resultados se expresaron como niveles de expresión de cada gen (una vez normalizados frente a los ARN de 18s como gen constitutivo) en cada condición experimental, respecto a su control correspondiente: 2 -ΔCt, donde Ct representa el ciclo umbral de PCR en el cual el programa detecta por primera vez un incremento apreciable de fluorescencia sobre la señal basal. Todas las determinaciones se realizaron por duplicado. (ΔCt = Ct (Gen de interés) – Ct (control endógeno 18S).
Análisis estadístico
Los resultados se expresaron como media +/- Desviación Estándar (DE). La comparación no paramétrica entre las medias de dos muestras se realizó por la prueba de Mann Whitney. Se utilizó ANOVA no paramétrica para comparar varias muestras (Kruskal-Wallis). Todos los valores con p<0,05 fueron considerados significativos.
Resultados
En primer lugar, se llevó a cabo la caracterización de las ASCs procedentes del cultivo primario de los lipoaspirados. El análisis fenotípico de ASC por citometría de flujo reveló que 99,6% eran CD90+, 91,6% CD44+, 90,4% CD29+, 4,2% CD34+, 2,2% CD45+. Además, se analizaron los marcadores CD106, CD155, KDR y CD31, todos ellos de expresión negativa para ASCs (Tabla 1).
Tabla 1 Caracterización por citometría de flujo de las ASCs aisladas de tejido adiposo obtenido por lipoaspirado

Para evaluar el potencial de diferenciación osteogénico de ASC, se las sometió a diferenciación osteoblástica in vitro. Como se muestra en la Figura 1A, la actividad ALP reveló que las ASCs en medio de diferenciación osteogénica habían sufrido dicha diferenciación. La tinción citoquímica rojo de alizarina (Figura 1B) también reveló el resultado positivo de la diferenciación osteogénica.

Figura 1 Respuesta del cultivo primario de ASC a los medios específicos de diferenciación de linaje celular(A) Detección de la actividad fosfatasa alcalina (ALP) y (B) detección de la diferenciación osteogénica de 14 días mediante la tinción con rojo de alizarina para detectar mineralización
Se evaluó la expresión génica de algunos de los marcadores de diferenciación osteogénica como OPG y Runx2 [12,13,24]. Se observó que la expresión de estos dos marcadores analizados aumentó significativamente (p<0,05) tras 14 días de cultivo con el medio de diferenciación osteogénica (Figura 2A-B).

Figura 2 Cambios en la expresión génica (analizada por PCR a tiempo real) de factores de diferenciación ósea: (A) OPG, (B) Runx2, a distintos tiempos de diferenciación osteoblástica(*) p<0,05 vs. valor basal correspondiente.
Se evaluó la expresión génica del sistema VEGFA y su receptor inducido por la diferenciación osteogénica. Tal y como se puede observar en la figura 3, tras 7 días de diferenciación las células disminuyen significativamente la expresión del VEGF y de su receptor tipo 2.
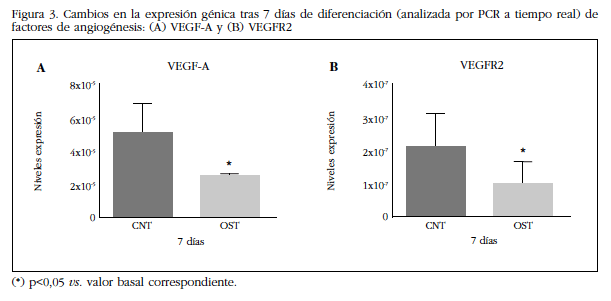
Figura 3 Cambios en la expresión génica tras 7 días de diferenciación (analizada por PCR a tiempo real) de factores de angiogénesis: (A) VEGF-A y (B) VEGFR2(*) p<0,05 vs. valor basal correspondiente.
Los ensayos de proliferación celular con medios suplementados con PTHrP y VEGF mostraron resultados diferentes. Se realizó un estudio de proliferación de las ASCs en respuesta a VEGF (160 pM) durante 24 horas en condiciones basales (medio de crecimiento) o con medio de diferenciación osteogénica.
Como se observa en la figura 4, el VEGF, aumentó de manera significativa la proliferación de las ASCs en medio de crecimiento, pero no tuvo efectos significativos sobre la proliferación de las células cultivadas en medio de diferenciación osteogénica (p<0,05). Los resultados indicaron que VEGF, aumentó la proliferación de las ASCs, pero siempre que éstas no hubieran iniciado su proceso de diferenciación. En respuesta a PTHrP(1-36) (100 nM) durante 24 horas, las ASCs no alteraron significativamente su índice de proliferación (Figura 4A). Cuando se realizó el ensayo de proliferación anteriormente descrito en medio de diferenciación osteogénica, se observó que éste cambiaba el fenotipo de las ASC, y alteraba su capacidad de respuesta ante estos factores. De forma que en este caso PTHrP incrementó el índice de proliferación en células ya comprometidas a la diferenciación osteoblástica (Figura 4B).

Discusión
En el presente estudio, ASCs de mujeres sanas, se expusieron a tratamientos cortos con PTHrP y VEGF en distintas condiciones de diferenciación y su capacidad proliferativa fue analizada. Nuestros resultados indican que las poblaciones más indiferenciadas serían más sensibles a VEGF mientras que los progenitores ya comprometidos hacia la diferenciación osteoblástica responden mejor a PTHrP(1-36), al mostrar una mayor expresión del receptor 1 de la PTH/PTHrP (PTH1R). Se utilizaron ASCs humanas procedentes de lipoaspirados como modelo in vitro de diferenciación osteogénica. Durante este estudio la caracterización de las ASCs se llevó a cabo mediante un estudio de los marcadores de superficie celular. Los datos obtenidos fueron los esperados según estudios previos [21,22]. Así mismo se estudió la capacidad de diferenciación osteoblástica de las ASCs. Dicha diferenciación se verificó por la actividad ALP de estas células tras 14 días en medio de diferenciación osteogénica. Del mismo modo, la tinción histoquímica de rojo de alizarina se utilizó para verificar dicha diferenciación complementando así el análisis de actividad ALP. Además, se evaluó la expresión génica de algunos marcadores como OPG y Runx2 [16,23]. Runx2 es esencial para la diferenciación osteoblástica, conduce a las ASCs hacia el linaje osteoblástico e inhibe su diferenciación hacia linaje adipogénico o condrogénico [23]. Además, Runx2 se ha descrito como un factor que mantiene a los osteoblastos en estado inmaduro sin diferenciarse a osteocitos [23]. Así mismo, estudios previos consideran a OPG como un marcador de diferenciación osteogénica temprana en células mesenquimales humanas [24]. Según lo esperado, nuestros datos muestran que la diferenciación osteogénica induce un aumento en la expresión de estos dos marcadores, OPG y Runx2, respecto al estado indiferenciado.
También se evaluó la expresión génica del sistema VEGFA y su receptor VEGFR2 en condiciones de diferenciación osteogénica. Tras siete días en presencia del medio de diferenciación osteogénica se observó una disminución en la expresión del VEGF y de su receptor tipo 2. En este estudio hipotetizamos que los efectos de VEGF y PTHrP sobre la proliferación celular de los osteoprogenitores son dependientes del estadio de diferenciación osteoblástica. Así, las poblaciones más indiferenciadas serían más sensibles a VEGF por su proximidad al nicho endotelial in vivo, mientras que los progenitores ya comprometidos hacia la diferenciación osteoblastica responderían mejor a PTHrP(1-36) al mostrar el receptor PTH1R un rol más importante en osteoblastos maduros y osteocitos. Tras confirmar su capacidad de diferenciación in vitro por expresión génica de Runx2 y mineralización, se analizó la respuesta proliferativa a estímulos con VEGF o PTHrP(1-36) de ASCs sometidas o no a inducción osteogénica. VEGF, pero no PTHrP(1-36), favoreció la capacidad proliferativa de las ASCs no inducidas, mientras que PTHrP(1-36), pero no VEGF, favoreció la proliferación de las ASCs previamente diferenciadas, confirmando el papel diferencial de estos factores de crecimiento en distintos estadios de diferenciación. Estudios previos indican que el VEGF secretado es crítico en la diferenciación de las BMSCs a osteoblastos, frenando su diferenciación a otros linajes celulares como el adipogénico [16,17]. Así mismo, Alonso y cols. [15] han mostrado como VEGF y PTHrP modulan la diferenciación y supervivencia de los osteoblastos. Nuestros resultados del estudio de proliferación de las ASCs en respuesta a VEGF durante 24 horas en medio de crecimiento o con medio de diferenciación osteogénica muestran un aumento en la proliferación de las ASCs crecidas en medio normal, pero sin efectos significativos sobre la proliferación de las células cultivadas en medio de diferenciación osteogénica, apoyando la noción de que VEGF ejerce un papel preponderante sobre la regulación de la proliferación en etapas tempranas de la diferenciación, aunque estudios previos apuntan también a una implicación de VEGF en la supervivencia de los osteoblastos [15].
Tras 24 horas de tratamiento con PTHrP(1-36), las ASCs indiferenciadas no vieron alterado su índice de proliferación. Sin embargo, en presencia de medio de diferenciación osteogénica, PTHrP(1-36) aumentó significativamente la proliferación, apuntando a la implicación de este factor en la proliferación de BMSCs comprometidas a la diferenciación osteogénica in vivo.
De esta forma podríamos concluir que, si bien estudios previos han mostrado que VEGF y PTHrP modulan la diferenciación y supervivencia de los osteoblastos [15], estos factores podrían regular la proliferación de los osteoprogenitores en función de su compromiso o no hacia la diferenciación osteoblástica, estando VEGF mas implicado en la proliferación de progenitores más indiferenciados y próximos al nicho perivascular, mientras que la PTHrP(1-36) estimularía células más comprometidas hacia la osteoblastogénesis.

