










 Custom services
Custom services
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
La pleitrofina (PTN) en una citoquina secretada por múltiples tejidos durante el desarrollo embrionario, y que en edades adultas se expresa de manera abundante en el cerebro y en el hueso1,2. La PTN está compuesta por 136 aminoácidos y su secuencia es muy rica en lisina y cisteína. Esta citoquina constituye junto a la midquina (Mdk), con quien comparte un 50% de homología, la familia de los factores de crecimiento y diferenciación de unión a heparina, teniendo ambas afinidad por la matriz extracelular ósea3-5. La PTN también es conocida como el factor estimulante de osteoblastos 1 (OSF-1) o factor de crecimiento unido a heparina (HB-GAM)6. Esta citoquina fue aislada inicialmente de los tejidos óseos y neuronales de ratas recién nacidas2,7,8 y posteriormente se han encontrado sus homólogos en muchas especies incluido el ser humano, siendo de un 90% la homología entre las distintas especies9,10.
Se ha descrito que la PTN ejerce sus efectos a través de su unión a los glucosamin-glucanos de varios receptores como el N-sindecano, también llamado sindecano 311, los sindecanos 1 y 412, la integrina αvβ313 y el receptor proteína tirosina fosfatasa beta/zeta (PTRPβ/ζ)14. También se ha postulado que la nucleolina pueda ser un receptor de baja afinidad de PTN15 y que la quinasa del linfoma anaplástico (ALK) puede tener alguna función en la señalización de PTN16.
Probablemente el receptor mejor estudiado de PTN ,y que además se ha demostrado que se expresa en osteoblastos17, es PTRPβ/ζ. Este receptor es una proteína tirosina fosfatasa cuya activación por parte de PTN produce su desestabilización y, por tanto, el cese en su actividad fosfatasa. Esto desencadena el aumento de fosforilación de sus sustratos (e.g. fyn-quinasa) dando lugar, entre otros efectos, a la activación del factor nuclear potenciador de las cadenas ligeras kappa de las células B activadas (NFκβ)18. Recientemente se ha confirmado la presencia de este receptor en el tejido óseo y su activación a través de PTN y de otro ligando, como es la proteína de unión 2 del factor de crecimiento similar a la insulina (IGFBP-2)19. Por otro lado, también se ha demostrado la expresión de N-sindecano en osteoblastos y su relación con la regeneración ósea6. En este caso la unión de PTN a N-sindecano produciría la fosforilación de src, lo que a su vez daría lugar a reorganizaciones en el citoesqueleto celular que permitirían un aumento de la migración celular20.
Entre sus las funciones más destacadas de la PTN, está su papel como promotor de la angiogénesis y de la migración de células endoteliales13,21, el crecimiento de las neuritas22 y su papel modulador de los procesos inflamatorios gobernados por la microglia en el sistema nervioso central23. Las funciones de PTN en el tejido óseo son diversas. Así, se ha descrito que PTN es capaz de inducir la proliferación de células osteblásticas de manera dependiente a su concentración y a la expresión de sus receptores24, promueve la diferenciación de células mesenquimales a condrocitos durante el desarrollo óseo25, y aumenta la migración y la adhesión a la matriz extracelular de células óseas6,26. De hecho, se ha investigado el papel de la PTN en relación con la masa ósea realizando experimentos de deleción o sobreexpresión del gen en modelos murinos. En el primer caso, se ha encontrado que la ausencia del gen no produjo un descenso en la masa ósea ni cambios significativos en las propiedades biomecánicas de estos huesos27. Estudios posteriores también encontraron que la estructura ósea de los ratones sin PTN no se veía alterada, pero que había un retraso en la maduración ósea en ratones de 2 meses de edad28.
También se ha estudiado del papel de la PTN en la mecanotransducción osteocitaria con diferentes resultados. En estudios in vitro se ha determinado que la carga mecánica conduce a una disminución de la expresión de PTN en las células óseas SaOs-2 y en osteoblastos primarios sometidos a este estímulo29. Sin embargo, Imai y cols. demostraron que células MLOY-4 (línea celular de osteocitos), tratadas con carga mecánica incrementaban su producción de PTN28. Estos datos están en consonancia con los hallados en un modelo animal de ratones hembra C57BL/6J sometidas a carga mecánica, en los que se produjo un aumento de la expresión de esta citoquina29. Además, en el mismo estudio se demostró que la ausencia de PTN (usando un ratón con el gen de la PTN delecionado) no influía en el aumento de masa ósea producido por la carga mecánica.
Por otro lado, los efectos de la sobreexpresión de la PTN en el tejido óseo sí que parecen tener un efecto protector en situaciones de pérdida de masa ósea, como la ingravidez. Los ratones transgénicos para la PTN están parcialmente protegidos frente a la pérdida de masa producida por el estado de ingravidez al ser sometidos a una estancia en la estación espacial internacional, relacionándose esta protección con un aumento de actividad osteoblástica30.
Dado que, como se ha comentado, la ausencia de PTN en hueso no parece influir de manera significativa en la estructura esquelética de los modelos murinos, en este trabajo nos propusimos averiguar cómo se alteraba la expresión de factores asociados con el mantenimiento correcto del metabolismo óseo en ausencia de PTN. Además, quisimos investigar cómo la expresión de estos genes estaba regulada ante una situación de inflamación aguda producida por inyección de LPS, tanto en presencia como en ausencia de PTN. Por último, comprobamos el efecto protector de la PTN en osteoblastos y osteocitos ante la presencia de un estímulo de muerte como es el etopósido.
MATERIAL Y MÉTODOS
Modelo animal
Se usaron ratones hembra de 3 meses deficientes en PTN (PTNKo) y con genotipo normal (WT) siendo el número de ratones empleados 9 para WT, 9 para PTNKo por grupo. Todos los ratones provenían de nuestro animalario, donde son criados de manera rutinaria. Para inducir un estado de inflamación aguda se inyectó por vía intraperitoneal una dosis de 7,5 mg/kg de lipopolisacárido 16 horas antes del sacrificio en 6 ratones WT y 5 ratones PTNKo. Los protocolos animales contaron con la aprobación del Comité de Bienestar Animal de la Universidad CEU-San Pablo y de la Consejería de Medio Ambiente de la Comunidad de Madrid, de acuerdo con el real decreto R.D. 53/2013 y la directriz europea 2010/63/EU.
Procesamiento de las muestras óseas y purificación del RNA
El RNA total fue extraído de las vértebras después de retirar los tejidos no óseos de las vértebras de las lumbares 1 a la 3, por medio de un pulverizador de tejido y por la disolución de este pulverizado en trizol (Invitrogen, Groningen, Holanda). El RNA fue extraído por medio de purificaciones por el método de clorformo:isoamílico (Sigma-Aldrich), seguido de precipitación con isopropanol, lavados posteriores con etanol al 70% y resuspensión en agua estéril libre de RNAasas. La retrotranscripción del ARN para obtener ADN complementario se llevó a cabo a partir de 2 μg de ARN usando el cDNA High capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, California, EE.UU.) en un termociclador Eppendorf mastercycler, siguiendo el siguiente protocolo secuencial: 10 minutos a 25ºC, 120 minutos a 37ºC y 5 minutos a 85ºC.
PCR cuantitativa
La PCR cuantitativa se realizó en un termociclador AB 7500 HD (Applied Biosystems). Usando sondas TaqMan MGB (Assay-by-DesignTM System, Applied Biosystems) para medir la expresión de los siguientes genes: osteoprotegerina (OPG), ligando del receptor activador para el factor nuclear κB (RANKL), osteocalcina (OCN), péptido relacionado con la parathormona (PTHrP), podoplanina (E11), conexina 43 (Cox43), el receptor del factor de crecimiento del endotelio vascular 2 (VEGFR2) y dickkopf-1 (Dkk1). Para producir la reacción de PCR se usó la polimerasa incluida en el kit Taqman mastermix (Applied Biosystems) aplicando el siguiente protocolo de 1 minuto a 95ºC, seguido de 40 ciclos de15 segundos a 95ºC y 1 minuto a 60ºC, recogiendo los datos de fluorescencia en cada paso de 60ºC. La expresión de del gen 18S ribosomal fue usado como gen de referencia para normalizar la expresión.
La variación de expresión de cada uno de los genes en los diversos grupos se calculó con relación a la expresión obtenida en los ratones WT siguiendo la siguiente fórmula:
Expresión gen de interés = 2-ΔΔCt, donde ΔΔCt = ΔCt tratamiento — ΔCt basal31,32. Todas las determinaciones se realizaron por duplicado.
Contaje de osteocitos
Las tibias de los ratones fueron también extraídas en el momento de sacrificio y conservadas en formalina 10% para su posterior procesamiento histológico. Una vez procesadas y teñidas con hematoxilina-eosina, se realizó el contaje de los osteocitos embebidos en las trabéculas en 2 a 4 cortes no seriados correspondientes a la región inmediatamente inferior a la placa epifisaria de cada una de las tibias. El número de osteocitos se normalizó por su área correspondiente de tejido óseo calculada por medio del programa Image J (ImageJ 15.3a, National Institutes of Health, EE.UU.), y siguiendo las recomendaciones de la American Society for Bone and Mineral Research33 . El número de osteocitos resultantes se calculó haciendo la media de cada contaje por ratón, y se expresó como la media ± error estándar de la media (EEM).
Cultivos de células in vitro y ensayo de viabilidad
Las células osteoblásticas murinas MC3T3-E1 se cultivaron en medio alfa MEM suplementado con un 10% de suero fetal bovino (FBS) (Sigma-Aldrich) y un 1% de penicilina-estreptomicina. La línea estable de osteocitos murinos MLOY-4 fue mantenida en placas de cultivo previamente colagenizadas y con medio alfa MEM al 2,5% de suero de ternero (Sigma-Aldrich), 2,5% de FBS y 1% de penicilina-estreptomicina. Ambas líneas celulares se incubaron a 17ºC en una atmósfera con 5% de CO2. Para los ensayos de viabilidad, ambas líneas celulares se incubaron en subconfluencia del 75% en placas de 6 pocillos y fueron pre-tratadas o no durante una hora con PTN (5,5 nM), y posteriormente incubadas en presencia de etopósido 50 μM (agente apoptótico) en 1% de suero fetal bovino durante 48 horas. A las 48 horas se contaron las células incluyendo las que había en el sobrenadante de los pocillos y el tripsinizado de cada pocillo, y se hizo un recuento celular con Trypan Blue 0,4% en PBS en la cámara de Neubauer, distinguiendo las células vivas de las muertas. Se calculó el porcentaje de células muertas de cada uno de los experimentos. Se realizaron 3 experimentos con cada condición por triplicado y se expresó el resultado como la media ± error estándar de la media (EEM).
Estadística
Los resultados se expresaron como media ± error estándar de la media (EEM). La comparación entre varios grupos se realizó mediante la prueba no paramétrica de Kruskall-Wallis con test a posteriori de U Mann-Whitney si procedía. Se consideró una p<0,05 como significativa. Los análisis se llevaron a cabo con el programa informático Graphpad InStat (San Diego, California, EE.UU.).
RESULTADOS
Expresión de genes en el tejido óseo en ausencia de PTN
En primer lugar, quisimos comprobar cómo variaba la expresión de genes relacionados con el metabolismo óseo en ausencia de PTN y en presencia de condiciones inflamatorias tras la administración de LPS. Entre la expresión de todos los genes analizados encontramos que la OCN estaba sobreexpresada de manera muy significativa en los ratones PTNKo (Figura 1A). Sin embargo, la expresión de un marcador de diferenciación ostecítico temprano como es la podoplanina (E11) no sufrió cambios significativos (Figura 1B). De la misma manera, no sufrieron cambios significativos en el ratón PTNKo la expresión de los moduladores de la actividad osteoblásticas y osteoclástica OPG y RANKL (Figuras 1C, 1D); el inhibidor de la vía Wnt, Dkk1 (Figura 1E); el VEGFR2 (Figura 1F); y los niveles de PTHrP (Figura 1G) y de Cox43 (Figura 1H). Por otro lado, cuando se trataron los ratones normales WT con LPS, encontramos que los niveles de OCN, Dkk-1 y VEGFR2 estaban significativamente disminuidos (Figuras 1A, 1D, 1F). Con respecto a los ratones PTNKo tratados con LPS encontramos un incremento significativo de la expresión de E11 (Figura 1B), PTHrP (Figura 1G) y Cox43 (Figura 1H) con respecto a los ratones WT tratados con LPS.
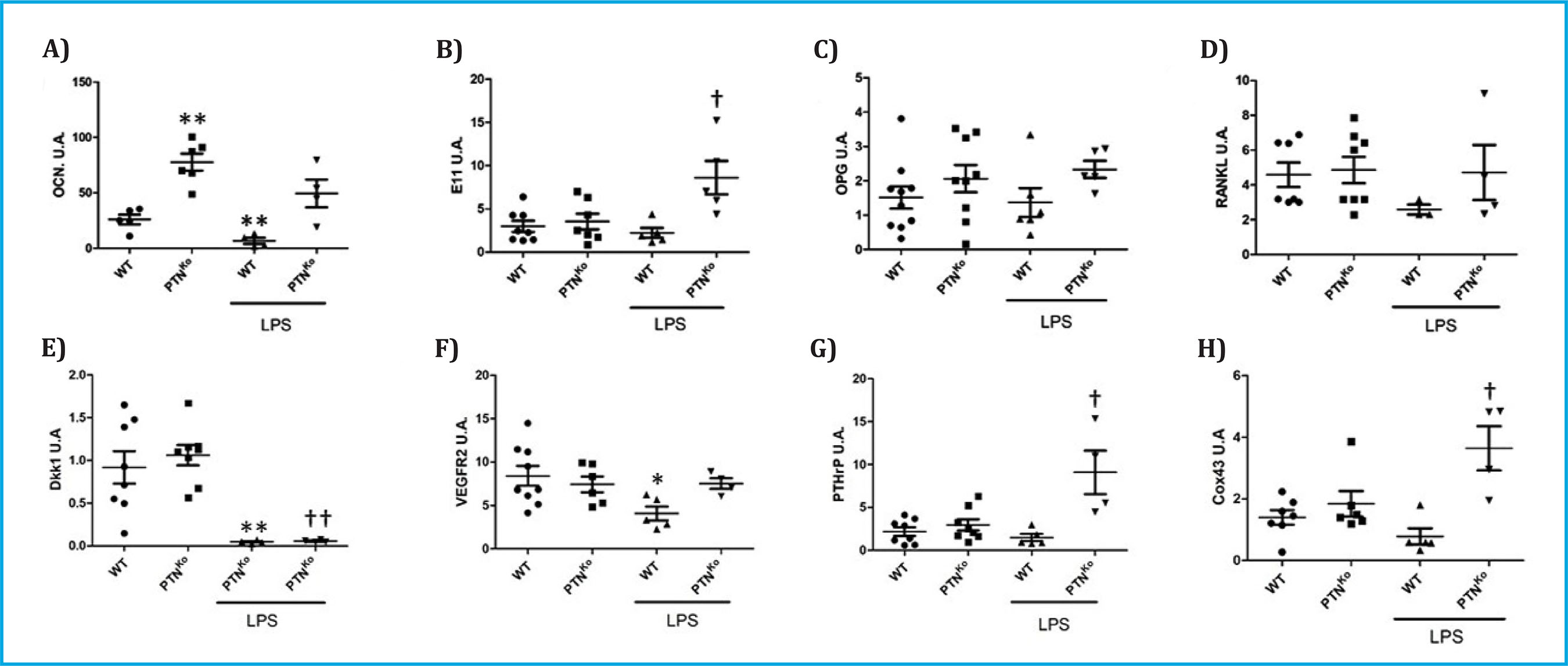
A) Osteocalcina (OCN) vs. WT, **p<0,05; B) Podoplanina (E11) vs. WT+LPS, †p<0,05; C) Osteoprotegerina (OPG); D) Ligando del receptor activador para el factor nuclear κB (RANKL); E) Dickoppf1 (Dkk1) vs. WT, **p<0,01; Dkk1 vs. WT+LPS, ††p<0,01; F) Receptor del factor de crecimiento del endotelio vascular 2 (VEGFR2) vs. WT, *p<0,05; G) Péptido relacionado con la parathormona (PTHrP) vs. WT+LPS, †p<0,05; H) Conexina43 (Cox43), vs. WT+LPS, †p<0,05. Los resultados de la qPCR se expresan con unidades arbitrarias (U.A.) una vez normalizados con el gen de control 18 S ribosomal.
Figura 1. Expresión de diferentes genes en las vértebras de ratones WT o PTNKO tratados o no con inyección de LPS
El número de osteocitos no sufrió cambios en ratones PTNKo, WT+LPS y PTNKo+LPS
Después de observar que el único gen cuya expresión estaba regulada era OCN, un gen relacionado con los estados muy avanzados de maduración osteoblástica34, y que esta estaba regulada al alza, pensamos que esta expresión podía reflejar un aumento del número de osteocitos en los ratones PTNKo (ampliamos también este contaje a los grupos WT y PTNKo tratados con LPS). Para ello teñimos las tibias aisladas de los ratones y realizamos un contaje exhaustivo de los osteocitos que se hallaban en las secciones de hueso tibial. Como se puede apreciar en las figuras 2A, 2B, el número de osteocitos fue similar en todos los grupos de ratones, no hallándose diferencias significativas en ningún caso.
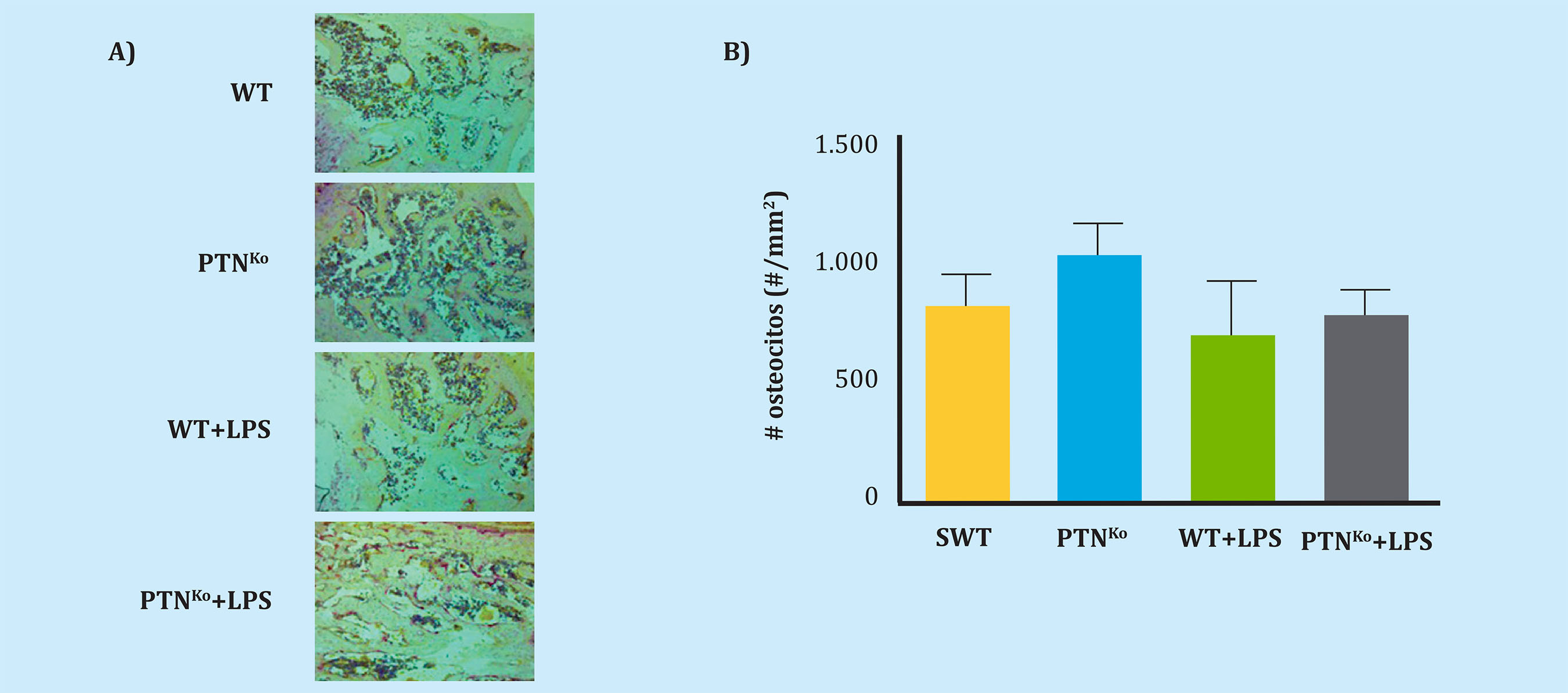
A) Imágenes representativas de las secciones de tibia donde se realizó el contaje de osteocitos debajo de la placa epifisaria de las tibias de los diferentes grupos experimentales; B) Cuantificación del número de osteocitos por mm2 de tejido óseo en los ratones WT, PTNKo, WT+LPS y PTNKo+LPS.
Figura 2. Número de osteocitos en la sección trabecular de las tibias de los ratones WT y PTNKo
Efecto protector de la PTN ante la inducción de muerte en células óseas
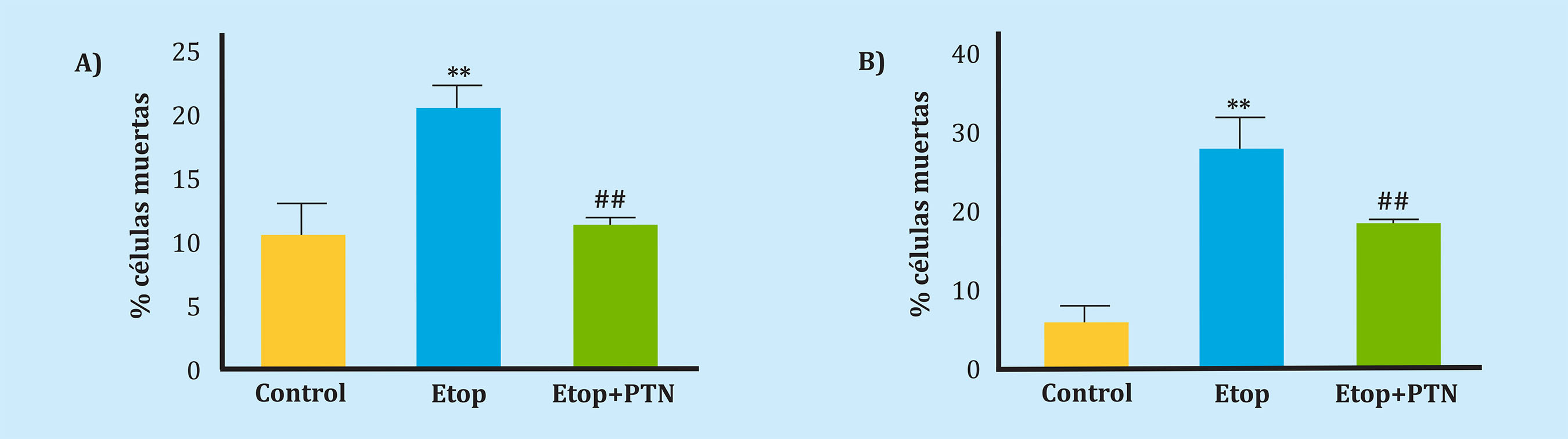
Dado que se ha descrito previamente el efecto proliferador de la PTN en osteoblastos MC3T324, quisimos comprobar el efecto protector de la PTN frente a la inducción de la muerte por etopósido en dos líneas de células murinas, una de osteoblastos (MC3T3) y otra de osteocitos (MLOY-4), in vitro. La administración de la PTN previamente al tratamiento con etopósido produjo un efecto protector, pues la muerte de células inducida por este agente se redujo significativamente en la línea MC3T3 de un 20,8% a un 11,5% y en la línea MLOY4 de un 27,5% a un 18,1% (Figuras 3A, 3B).
DISCUSIÓN
Es notorio el hecho de que la deleción de un gen tan importante para el desarrollo y mantenimiento del tejido óseo como la PTN no produzca un fenotipo acusado de alteraciones óseas. En primer lugar, se podría pensar que, ya que la PTN y Mdk tienen expresiones paralelas y funciones similares, la falta de PTN podría ser compensado por el aumento de Mdk. Sin embargo, esto no parece ser así, pues se ha demostrado que los ratones deficientes en Mdk tienen un incremento en la masa ósea29, apuntando a que en el tejido óseo ambas citoquinas no comparten las mismas funciones de manera total. En este trabajo quisimos comprobar cómo era la expresión de genes relacionados con el metabolismo óseo y si esta variación pudiera justificar, al menos en parte, la falta de efectos óseos en los ratones PTNKo.
Recientes investigaciones han desvelado que la OCN es una proteína que actúa regulando el correcto alineamiento de los cristales de hidroxiapatita con el colágeno, lo cual está directamente relacionado con la calidad ósea35, aunque también es usada como marcador de maduración osteoblástica36, y en estos momentos está bajo debate su papel como hormona producida por el hueso37-39. El hecho de que esté aumentada en los ratones PTNKo nos llevó a pensar que quizá el número de osteocitos de manera paralela podría estar incrementado en estos ratones, al ser un marcador de maduración tardía de los osteoblastos justo antes de convertirse en osteocitos, y pudiendo ser este uno de los mecanismos celulares de compensación de la falta de la PTN. Sin embargo, nuestros resultados (Figura 2A, 2B) demuestran que el número de osteocitos no varía significativamente en los ratones PTNKo comparados con los ratones WT ni tampoco en el resto de los grupos experimentales. Estos resultados concuerdan con los mostrados por Lehman y cols., en los cuales se observó un aumento de las expresión de OCN en cultivos primarios procedentes de ratones PTNKo y con un número equivalente de osteocitos entre los ratones normales y los PTNKo27. Cabe pensar que es probable que el aumento de osteocalcina tenga otros efectos en hueso y que la matriz ósea de estos ratones tenga una composición alterada. Para comprobar este extremo serían necesarios estudios posteriores de resonancia magnética nuclear donde se analice la calidad y composición del hueso de los ratones PTNKo.
Por otra parte, y dada la importancia que tiene PTN en determinados procesos inflamatorios, quisimos también investigar cuál era su papel en la expresión de estos genes ante una inflamación aguda provocada por inyección de LPS. En este caso descubrimos que la expresión de E11, PTHrP y Cox43 estaba incrementada en los ratones PTNKo frente a los ratones WT tratados con LPS. La PTHrP es una citoquina que modula el remodelado óseo asemejando de manera local las acciones de la parathormona40. Esta citoquina está implicada en los procesos inflamatorios que se dan en la médula ósea, pues promueve la expresión del factor inflamatorios MCP-1 en las células endoteliales y osteoblastos41. E11 gobierna los primeros pasos de la transición osteoblasto-osteocito42, siendo su sobreexpresión un marcador de tumores óseos43 y de inflamación en otros tejidos y diversos tumores44. Por otro lado, la Cox43 forma parte de las comunicaciones que establecen los osteocitos entre ellos para formar la red osteocitaria. El papel de las conexinas en los procesos inflamatorios es diverso y variable en función de los distintos tejidos45. Se han descrito incrementos de expresión de Cox43 que están relacionados con enfermedades como la artrosis46, y una mayor expresión en sinoviocitos ante estímulos como el LPS47. Parece ser que la disminución de los niveles de Cox43 estaría relacionado con una disminución de los procesos inflamatorios en los procesos de artritis47. En nuestro estudio, la ausencia de PTN, al menos en el compartimento óseo, tendría un papel perjudicial desde un punto de vista inflamatorio, pues permitiría la sobreexpresión de los tres genes citados (Figuras 1B, 1G, 1H) al contrario de lo que ocurre en el ratón WT tratado con LPS, y, por tanto, daría lugar a efectos indeseados. Evidentemente, la verificación última de estas observaciones requiere posteriores estudios tanto in vivo como in vitro para desentrañar los mecanismos moleculares desencadenados por la ausencia de PTN en este contexto.
Una de las limitaciones de este estudio es que la edad de los ratones era de 3 meses, un poco alejado de los 5 meses en que se puede considerar un ratón maduro desde el punto de vista esquelético. Hay que tener en cuenta que los modelos murinos no sufren cierre de las placas de crecimiento ni depleción estrogénica, lo cual hace que siempre tengan un pequeño crecimiento óseo a lo largo de su vida. A este respecto, hay que señalar que se no se han descrito en ratones deficientes para PTN de 50 semanas graves defectos óseos27, aunque otros autores si han encontrado pequeños retrasos del crecimiento en ratones de 4 meses28. Además, en este último estudio, se ha demostrado que las características biomecánicas de los huesos largos están alteradas al tener disminuidos los parámetros de rigidez, resistencia a la ruptura, y dureza óseas28. Dado que la PTN contribuye a la correcta alineación de los cristales de hidroxiapatita, futuros estudios en los huesos de ratones adultos o, incluso, envejecidos con ausencia de PTN, deberían incluir análisis de estos huesos por medio de resonancia magnética nuclear para determinar si la ausencia de PTN puede provocar la creación de un hueso frágil en edades avanzadas. Otra de las limitaciones del estudio, es que, dado que no se han incluido zonas corticales al contar los osteocitos, no podemos establecer si existen diferencias entre los compartimentos cortical y esponjoso, por lo que este tipo de análisis debe de ser realizado en estudios posteriores.
Es de destacar que se ha comprobado previamente, que la administración de la PTN in vitro promueve la proliferación y la diferenciación de células osteoblásticas24. Por ello, quisimos comprobar, hasta donde sabemos por primera vez, el efecto protector de la PTN frente a un estímulo pro-apoptótico como es el etopósido en células osteblásticas (MC3T3) y osteocitos (MLOY4), observando que la PTN tiene una acción protectora. Este resultado apunta a un posible papel adicional de la PTN en situaciones de fractura o inflamación crónica en las cuales podría proteger, al menos parcialmente, a las células óseas. Es cierto que en este estudio no se ha comprobado el papel de protección que pueda desempeñar la PTN en cuanto a la muerte medida por LPS. En este sentido, estudios previos han demostrado que este agente produce un fenotipo senescente en los osteocitos y un aumento de la resorción ósea mediada por la producción del IL-6 y RANKL por los osteocitos, más que una muerte en un corto periodo de tiempo, como es el tiempo de administración de LPS que se ha realizado en este estudio. Sin embargo, sería interesante estudiar el papel de la PTN en estos contextos en estudios posteriores y en modelos adecuados48,49.
Por todo lo anterior, podemos concluir que la deficiencia de PTN está acompañada de un aumento de la expresión de OCN, y que la inducción de inflamación aguda por medio de LPS en ratones deficientes para PTN produce la sobreexpresión de E11, PTHrP, y Cox43. Así mismo, hemos demostrado el papel protector de la PTN en células osteblásticas y en osteocitos ante un estímulo de muerte celular.

