










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista de Diagnóstico Biológico
versión impresa ISSN 0034-7973
Rev Diagn Biol vol.51 no.3 jul./sep. 2002
REVISIONES
La citometría de flujo en el análisis funcional de las plaquetas.
II. Aplicaciones clínicas
María-do-Céu Monteiro*, Marcial Martínez**, José Enrique O'Connor***
*Departamento de Bioquímica, Instituto Politécnico de Saúde-Norte, Gândra, Portugal.
**Unidad de Citometría de Flujo, Departamento de Biopatología Clínica, Hospital Universitario La Fe, Valencia.
***Centro de Citometría, Departamento de Bioquímica y Biología Molecular, Facultad de Medicina y Odontología,
Universidad de Valencia, España.
Palabras Clave: Activación, plaqueta, agregación, glicoproteinas, calcio, citometría de flujo.
Key words: Activation, platelet, aggregation, glycoproteins, calcium, flow cytometry.
Recibido: 10-I-02
Aceptado: 5-VI-02
Correspondencia: Dra. María-do-Céu Monteiro
Departamento de Bioquímica, Instituto Politécnico de Saúde-Norte
Rua Central de Gândra 1317, 4585-116 Gândra PRD, Portugal
Subvenciones: Ayuda PRODEP, Ministério de Eucaçâo, Portugal para la realización del Doctorado de María-do-Céu Monteiro. Convenio Universidad de Valencia-Izasa para la Creación y Mantenimiento del Centro de Citometría en la Universidad de Valencia. Primer Premio de Laboratorios LabMed, Portugal.
Introducción
Las alteraciones en la función plaquetaria están asociadas a condiciones patológicas que llevan tanto a disfunciones tromboembólicas como hemorrágicas. Una activación plaquetaria excesiva causa trombosis, mientras que la activación insuficiente causa hemorragia. Por esta razón, los estudios de activación y reactividad plaquetaria son relevantes en el conocimiento de la fisiopatología de la hemostasia y en las estrategias terapéuticas.
La funcionalidad plaquetaria puede ser evaluada por variadas metodologías que detectan alteraciones morfológicas, bioquímicas y de la composición de la membrana, asociadas a la activación celular. Entre los principales métodos utilizados para analizar la función plaquetaria se cuentan los métodos basados en la agregación plaquetaria, los basados en la liberación de productos plaquetarios, los métodos basados en el análisis microscópico y el análisis por citometría de flujo.
El análisis de las propiedades funcionales de las plaquetas ex vivo, utilizando las técnicas convencionales, como la agregación plaquetaria y la cuantificación de productos liberados por las plaquetas, refleja el comportamiento de la población plaquetaria total en la muestra en estudio. Sin embargo, diferentes poblaciones en la muestra pueden contribuir de forma diferente al resultado global y no es posible discriminarlas. En cambio, la citometría de flujo proporciona un análisis multiparamétrico de células individuales, constituyendo por ello una técnica extremadamente útil en el estudio de las plaquetas. Hoy es evidente que el análisis de las plaquetas por citometría de flujo es un área de investigación de gran interés y con innumerables aplicaciones clínicas.
Descripción resumida de la técnica
La citometría de flujo es una técnica con gran potencial analítico, que reside principalmente en la posibilidad de medir diversos parámetros en decenas de millares de células individuales en pocos segundos. Básicamente el citómetro de flujo mide propiedades de absorción y dispersión de luz por la célula o partícula subcelular, y fluorescencia emitida por fluorocromos unidos a componentes celulares de interés, inducidas por una iluminación apropiada .
Los componentes básicos de un citómetro de flujo están esquematizados en la Figura 1 e incluyen:
1. Fuente de luz, que en general es un rayo láser, por tratarse de una fuente de luz altamente direccionada, monocromática, coherente e intensa. La luz emitida, además, está concentrada en un haz estrecho. La mayoría de los citómetros actuales disponen de un láser de ión argón.
2. Cámara de flujo, en la que se orienta el flujo de células y se consigue el paso de éstas en forma individual.
3. Sistema óptico, constituido por lentes, espejos dicroicos y filtros, que enfocan el haz de luz para iluminar la muestra y descomponen la emisión de fluorescencia en sus componentes individuales.
4. Fotomultiplicadores y convertidores analógicos digitales, que detectan y convierten las señales luminosas en impulsos eléctricos y estos en señales digitales.
5. Ordenador para adquisición, almacenamiento y análisis de los datos en tiempo real o en modo de matriz de datos.
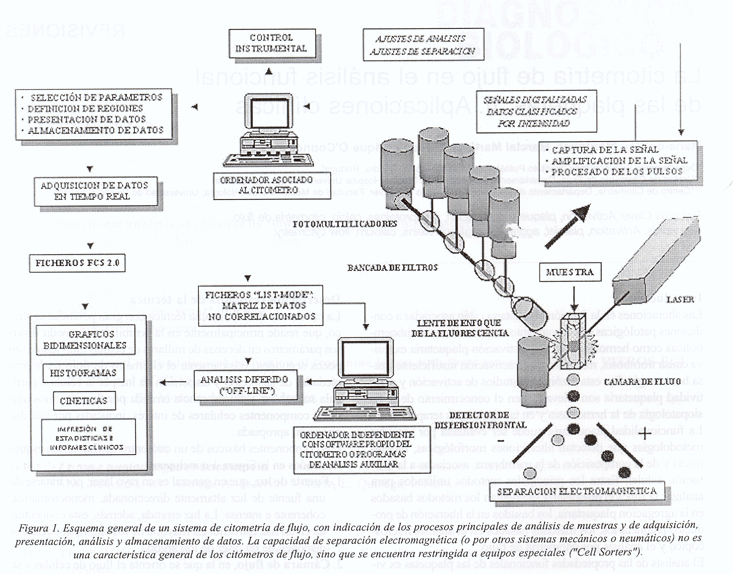
Las partículas a analizar (células, microorganismos, cromosomas, orgánulos celulares o partículas inertes) deben constituir una suspensión y son introducidas en un sistema hidrodinámico, que crea un flujo laminar, de modo que las células son forzadas, por una capa de líquido envolvente ("sheath fluid") a fluir en una fila única en el centro de la corriente, lo que se denomina "enfoque hidrodinámico". Las partículas transportadas por el flujo de líquido interceptan el haz de luz causando una dispersión de la luz incidente. La mayor o menor dispersión de luz según diferentes ángulos permite cuantificar el tamaño relativo y la complejidad (morfología de la superficie, granulosidad interna) de la célula. A pesar de que la luz se dispersa en todas las direcciones, la cantidad de luz dispersa en ángulos pequeños hasta 10-20° ("Forward Scatter", FS), alrededor del eje del haz de láser, está relacionada con el tamaño relativo de la célula (cuanto mayor es la partícula, mayor la dispersión). Las plaquetas, por ejemplo, tienen valores de FS inferiores a cualquier otra célula de la sangre, debido a su pequeño tamaño, lo que normalmente permite su distinción. Sin embargo, la correlación entre el tamaño de la célula y el valor de FS está afectada por diversos factores, concretamente por la simetría de la célula, la viabilidad y la presencia de componentes celulares que absorben la luz incidente. Este aspecto debe ser considerado cuando se usa la medición de FS para estimar tamaños relativos de células, en poblaciones de células heterogéneas.
La intensidad de la luz del láser que se dispersa ortogonalmente en relación al haz de luz ("Side Scatter", SS), representa principalmente la luz reflejada por estructuras internas o de la superficie de la célula y es interpretada como un índice de granularidad. La distinción entre diferentes tipos de células de sangre humana es más fiable cuando se utiliza la combinación biparamétrica de FS y de SS, que con cada uno de los parámetros solos (Figura 2).

La utilización de fluorocromos permite detectar y cuantificar un gran número de parámetros celulares. La fluorescencia emitida por estas moléculas, cuando son excitadas por el haz de luz, genera información acerca de su presencia en células o partículas individuales. La determinación de la fluorescencia permite una detección sensible de diferencias cuantitativas entre células o partículas. Por ejemplo, diferencias menores de 2.000 moléculas de fluorescencia por célula o partícula pueden ser detectadas con los fluorocromos más comunes.
Al igual que la dispersión de luz, la fluorescencia se emite en todas las direcciones, pero es monitorizada por un detector óptico en posición ortogonal con respecto al haz de iluminación (Figura 1). Pueden ser cuantificadas, separadamente, emisiones de varios fluorocromos diferentes en la misma célula, usando espejos dicroicos, que son separadores del haz luminoso, y filtros que seleccionan longitudes de onda adecuadas.
El análisis multiparamétrico de células individuales permite que sean discriminadas subpoblaciones de células. Durante la adquisición de los datos, es posible definir ventanas electrónicas ("gates"), que permiten cuantificar otros parámetros solo sobre la subpoblación de interés.
En el análisis de la fluorescencia emitida por fluorocromos añadidos a las células, debe tenerse en cuenta la fluorescencia celular intrínseca o autofluorescencia, que es causada, principalmente, por nucleótidos de pirimidina y flavina. Estas moléculas confieren fluorescencia azul o verde de baja intensidad a las células cuando son excitadas por luz ultravioleta o azul. La autofluorescencia también puede ser usada como un parámetro discriminador: por ejemplo, la fluorescencia emitida por moléculas tales como NADH y FMN/FAD, proporciona un marcador del estado de oxidación-reducción de las células.
En la interpretación y en la validación de los datos de la citometría de flujo son de particular importancia la linealidad y la estabilidad del aparato. La inestabilidad y/o irreproductibilidad pueden resultar de fluctuaciones en la potencia del láser o en el flujo de muestra y el desalineamiento de los sistemas de lentes. La no linearidad puede surgir de limitaciones en la linearidad de respuestas del fotomultiplicador o defectos en otros procesadores de señales electrónicos, y de la extinción ("quenching") dependiente de la concentración de los fluorocromos. La linearidad del aparato puede, con todo, ser evaluada usando microesferas fluorescentes u otras partículas de referencia.
Una característica particularmente útil de algunos citómetros de flujo es la posibilidad de separar físicamente las células (cell sorting) de interés, de muestras celulares heterogéneas.
Parámetros plaquetarios susceptibles de ser analizados por citometría de flujo
Morfología
En el análisis citométrico las señales de dispersión frontal (FS) y lateral (SS) de luz dependen de la morfología celular, estando FS relacionada con el tamaño y SS con la complejidad de la textura celular. Así, durante la activación de la plaqueta, la reorganización del citoesqueleto determina la pérdida de la forma discoide. La plaqueta se torna esferoide, emite pseudópodos y se desgranula. Estos cambios morfológicos están asociados a cambios concomitantes de las señales de FS y SS (Figura 3).

Antígenos plaquetarios
La utilización de anticuerpos conjugados con moléculas fluorescentes ha permitido la detección y cuantificación de numerosos antígenos plaquetarios por citometría de flujo. En contraposición a la tinción de componentes celulares con diversos colorantes fluorescentes, la tinción con anticuerpos (inmunofenotipaje) proporciona un análisis de alta especificidad y sensibilidad. Para la mayoría de las glicoproteinas de la membrana plaquetaria se conoce su función y existen anticuerpos monoclonales disponibles. Algunos antígenos se expresan en la superficie tras la activación plaquetaria, pero no en la plaqueta en reposo.
Una de las utilidades del inmunofenotipaje de plaquetas es determinar inequívocamente que las partículas en estudio son plaquetas. Marcadores como CD41, CD42 y CD61, específicos para glicoproteinas constitutivas de la membrana plaquetaria, permiten distinguir las plaquetas del ruido de fondo o "debris" (Figura 4). Adicionalmente, es posible caracterizar anomalías en la expresión de glicoproteinas de la membrana, como ocurre en el síndrome de Bernard Soulier y en la trombastenia de Glanzmann.

Procesos de activación
Durante la activación de la plaqueta ocurren diversas alteraciones a nivel de los lípidos y de las proteínas de la membrana plaquetaria. Estas pueden ser detectadas por citometría de flujo, utilizando anticuerpos monoclonales específicos conjugados con un fluorocromo, como por ejemplo la fluoresceína o la ficoeritrina. Los cambios de la membrana incluyen a) Alteraciones cuantitativas de glicoproteínas componentes de la membrana en reposo; b) Alteraciones conformacionales de receptores y sus ligandos; c) Exposición de componentes de la membrana de gránulos plaquetarios; d) Unión a la membrana de las plaquetas de proteínas solubles liberadas durante la secreción; e) Expresión de actividad procoagulante y formación de micropartículas; f) Otros fenómenos bioquímicos intracelulares de activación, como flujos iónicos, formación de peróxidos y radicales libres o cambios de potencial de membrana.
A) Alteraciones cuantitativas de glicoproteínas componentes de la membrana en reposo:
Se han realizado diversos estudios usando anticuerpos para el complejo GPIIb/IIIa, tales como 7E3, LJ-CP3 y LJ-P9, AP-3, en los que se ha demostrado un aumento de su unión después de la activación de la plaqueta. Estos anticuerpos se unen tanto a la plaqueta activada como en reposo, y el aumento en los lugares de unión tras la activación es debido al desplazamiento de moléculas de GPIIb/IIIa de los gránulos a hacia la superficie de la membrana durante el proceso de secreción1,2.
Al contrario de GPIIb/IIIa y GPIV, cuya expresión aumenta tras la activación de la plaqueta, la unión a la superficie de la plaqueta de anticuerpos monoclonales para diferentes epítopos del complejo GPIb/IX/V disminuye. Estudios realizados en sangre entera demuestran que la trombina induce una disminución acentuada de la unión de los anticuerpos monoclonales 6D1, específico para el factor de Von Willebrand ( vWF) ligado a GPIb; FMC25, específico para GPIX y AK1 específico para el complejo. Otros agonistas, tales como el ADP, adrenalina y colágeno ejercen un efecto similar sobre la expresión de GPIb/IX/V, pero en menor grado3,4.
B) Estados conformacionales de GPIIb/IIIa y de sus ligandos:
El anticuerpo PAC-1 se dirige al lugar de unión del fibrinógeno en el complejo GPIIb/IIIa. Este lugar de unión no está expuesto en las plaquetas en reposo. La activación de las plaquetas por agonistas (ADP, adrenalina y trombina), resulta en alteraciones conformacionales de GPIIb/IIIa, que exponen el lugar de unión del fibrinógeno. Así, PAC1 solamente se une a plaquetas activadas, permitiendo detectar la conversión del complejo GPIIb/IIIa en un receptor funcional para el fibrinógeno5-7. Esta alteración conformacional se ha determinado también por transferencia de fluorescencia (FRET, Fluorescence Resonance Energy Transfer) entre fluoróforos dadores y receptores unidos a anticuerpos anti-GPIIb y anti-GPIIIa8. Cuando se compara este fenómeno con la secreción de serotonina y PF4, no se observa una relación directa, lo que sugiere que la unión del PAC-1 puede ser independiente de la secreción1.
Después de la activación del complejo GPIIb/IIIa, éste se une a moléculas de adhesión (fibrinógeno, vWF, vitronectina y fibronectina) y esta unión puede ser monitorizada con anticuerpos específicos para cada uno de los ligandos9. Algunos anticuerpos monoclonales reconocen preferencialmente el ligando acomplejado10. Estos sitios de unión para los anticuerpos se designan genéricamente por "RIBS" (Receptor-Induced Binding Sites). Existen, además, anticuerpos que reconocen alteraciones en el receptor inducidas por la interacción con el ligando11. Los anticuerpos contra tales "LIBS" (Ligand-Induced Bindig Site) reconocen epítopos en GP IIb/IIIa, que son expresados cuando un ligando que contiene la secuencia Arg-Gly-Asp se une al receptor. Por ejemplo, el anticuerpo monoclonal D3 reconoce un lugar de unión LIBS en GPIIIa, cuya expresión es inducida por fibrinógeno y vWF12,13. El anticuerpo monoclonal PMI-1, reconoce un epítopo de GPIIb/IIIa, que es expuesto después de la unión del fibrinógeno al complejo. La utilización del anticuerpo PMI-1 puede ser más ventajosa que PAC-1 para detectar alteraciones conformacionales de GPIIb/IIIa y la consiguiente agregación, toda vez que la unión del fibrinógeno no bloquea el lugar de unión de este anticuerpo7.
Otro aspecto importante de la aplicación de esta técnica, es la posibilidad de analizar diversas etapas de la secuencia fisiológica de la activación. Por ejemplo, con los anticuerpos monoclonales actualmente disponibles, se puede evaluar secuencialmente el proceso de exposición de lugares receptores para el fibrinógeno en el complejo GPIIb/IIIa, la unión del fibrinógeno, las alteraciones inducidas por esta unión tanto en el complejo como en la molécula de fibrinógeno y, finalmente, la agregación.
C) Expresión de componentes de la membrana de los gránulos plaquetarios:
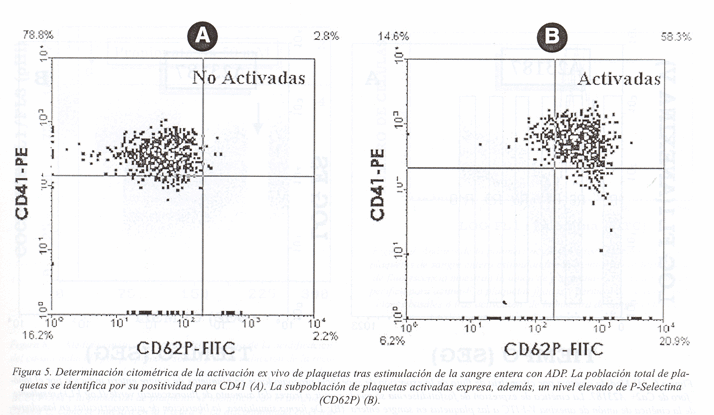
Otra alteración que ocurre en la superficie de la membrana durante la activación es la fusión de las membranas de los gránulos a con la membrana plasmática. Este proceso puede ser detectado con los anticuerpos S12 y KC4, que reconocen la glicoproteína de la membrana de los gránulos a denonimada P-selectina (CD62P)14. CD62P es la glicoproteína de la membrana de los gránulos mejor caracterizada y se han obtenido diversos anticuerpos monoclonales específicos para esta proteína, muchos de los cuáles inhiben sus funciones adhesivas15-17 (Figura 5). Más recientemente, se ha identificado y caracterizado otra proteína asociada a la membrana de los gránulos a, denominada GMP-33. El uso del anticuerpo monoclonal RUU-SP 1.77, permitió detectar un aumento de la expresión de esta proteína en la superficie de la plaqueta después de la activación con trombina18.
La membrana de los lisosomas contiene la glicoproteína GP53, conocida también como CD63 o proteína integral de la membrana lisosomal (LIMP), y dos proteínas homólogas referidas como LAMP-1 y LAMP-2 (Lysosome-Associated Membrane Proteins), que se expresan en la superficie de la membrana después de la activación. El anticuerpo específico 2.28 permite detectar la expresión de CD63 tras la estimulación de la célula con trombina18,19.
D) Expresión de proteínas solubles liberadas durante la secreción:
Tras la activación, diversas proteínas de los gránulos a liberadas al medio extracelular, pueden unirse a la superficie de la membrana en cantidad suficiente para ser detectadas por anticuerpos monoclonales apropiados. Estas proteínas incluyen las cuatro proteínas adhesivas que ligan GPIIb/IIIa, es decir, fibrinógeno, vWF, fibronectina y vitronectina; la trombospondina y el PF420-23.
E) Expresión de una superficie procoagulante:
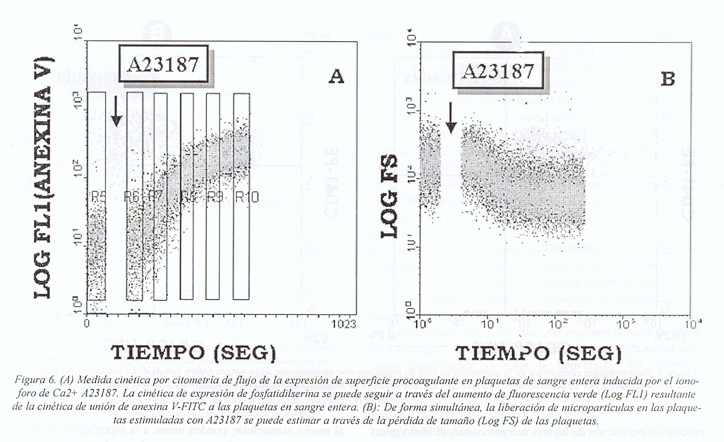
Las plaquetas activadas proporcionan una superficie procoagulante que permite la activación del factor X y de la protrombina. La aparición de lugares de unión los factores de la coagulación está asociada a la exposición de fosfatidilserina en la capa externa de la membrana plasmática y a la formación de pequeñas vesículas membranares (micropartículas) a partir de la superficie de la membrana.
Las micropartículas pueden distinguirse de las plaquetas por citometría de flujo, combinando la utilización de un discriminador de tamaño y un marcador para un antígeno específico de la plaqueta. La unión de los factores VIIIa y Va a las plaquetas y micropartículas puede ser detectada usando anticuerpos monoclonales específicos para estos factores. Para la detección del factor Va se ha utilizado el anticuerpo V237 y para el factor VIIIa el anticuerpo 1B37,24.
La expresión de fosfatidilserina, tanto en las micropartículas como en las plaquetas activadas, puede ser detectada directamente por la unión de anexina-V conjugada con un fluorocromo (Fig. 6). Esta proteína, en presencia de concentraciones fisiológicas de calcio, se une específicamente y con gran afinidad a los fosfolípidos aniónicos, que pasan de la capa interna a la capa externa de la membrana, tras la activación plaquetaria25,26.
F) Parámetros intracelulares bioquímicos de activación:
En las respuestas bioquímicas que median la activación están implicadas numerosas alteraciones intracelulares detectables y cuantificables por citometría de flujo. La aplicación de la citometría de flujo al estudio de estos mecanismos de transducción de la señal está limitada por la posibilidad de marcar con un colorante fluorescente específico las moléculas a analizar. Una gran variedad de colorantes pueden reaccionar directamente con moléculas intraplaquetarias o pueden ligarse a ellos dependiendo del estado funcional de la plaqueta. Por otra parte, la capacidad multiparamétrica de la citometría de flujo permite analizar simultáneamente dos o más eventos, cuando se utilizan fluorocromos que emiten a diferentes longitudes de onda.
La activación inducida por la mayoría de los agonistas plaquetarios produce un incremento del Ca2+ libre debido tanto al influjo a través de la membrana, como a la liberación de los depósitos intracelulares27. La citometría de flujo proporciona un método conveniente para evaluar las cinéticas de liberación de Ca2+ intracelular cuando se utilizan indicadores sensibles que pueden ser incorporados por las células intactas. Los fluorocromos Indo-1, Fura-2 y Fluo-3 se han sido utilizado para monitorizar el proceso de señalización por Ca2+ en preparaciones de plaquetas lavadas o filtradas en gel28-31. Mas recientemente se ha descrito un método para analizar la movilización del Ca2+ intraplaquetario directamente en plaquetas de sangre entera, utilizando Fluo-3 y CD4132 (Figura7).

La cinética de las variaciones de Ca2+ intraplaquetario inducidas por diferentes agonistas, se han analizado por citometría de flujo y por espectrofluorimetría. Mientras que mediante la espectrofluorimetría se obtiene un valor promedio de la respuesta de todas las células de la muestra analizada, la citometría de flujo permite clasificar la población según las respuestas en células individuales y distinguir subpoblaciones que responden de modo diferente. Así, se ha demostrado que algunas células presentaban respuesta máxima a 50% de la dosis saturante de trombina, mientras que otras no respondían completamente. Esta fracción no estimulada era, con todo, capaz de responder a dosis más elevadas de trombina29,33.
Los fluorocromos BCECF34 y SNARF-135 ofrecen la posibilidad de medir el pH intracelular. Las células mantienen su pH intracelular (pHi) en un estrecho rango de valores. Sin embargo, se producen cambios en el pHi en respuesta a la señalización celular. Las mencionadas sondas de pH son ácidos débiles, con pKs próximos a 7,0 y sus formas protonadas tienen diferentes espectros de emisión de la base conjugada. Midiendo el cociente de las dos emisiones resultantes se obtiene una señal proporcional al pHi e independiente del contenido intracelular del fluorocromo (Fig. 8). Las formas acetoximetil ester (AM) de los fluorocromos penetran en las células intactas y son hidrolizadas por esterasas inespecíficas intracelulares para generar el fluorocromo libre, retenido en el interior celular33. En diversos estudios de espectrofluorimetría, utilizando BCECF se observó un incremento del pHi en plaquetas estimuladas con trombina36-38.

La polimerización de la actina globular (G-actina) en actina filamentosa (F-actina) es uno de los acontecimientos más tempranos de la activación de la plaqueta y está implicada en el cambio de forma, la centralización de los gránulos y la retracción del coágulo. La F-actina puede ser detectada utilizando como sondas la falacidina o faloidina fluorescentes (Figura 9). Siguiendo esta aproximación, Oda et al.39 lograron determinar el contenido en F-actina, por citometría de flujo, en plaquetas mantenidas a diferentes temperaturas, estimuladas con ADP y adrenalina, y en presencia de citocalasina, un inhibidor de la polimerización de actina. En paralelo, efectuaron mediciones de la concentración de Ca2+ citosólico y evaluaron la expresión de P-selectina.

La mepacrina ha sido utilizada en algunos estudios como indicativa de tinción y secreción de los gránulos densos40.
Otros marcadores intracelulares potenciales de activación plaquetaria, para los cuáles no hay referencias de su estudio por citometría de flujo, son los cambios de potencial de membrana citoplasmática41,42 y mitocondrial, y la producción de radicales libres y peróxido de hidrogeno43-45.
Análisis citométrico del proceso de trombopoyesis
En condiciones caracterizadas por un elevado recambio de plaquetas, como en la púrpura trombocitopénica idiopática, se observa un incremento de plaquetas reticuladas en circulación. Las plaquetas reticuladas, por analogía con los reticulocitos eritroides, representan las subpoblación plaquetaria más joven. Estas plaquetas recién liberadas de la medula ósea siguen conteniendo RNA y pueden ser distinguidas de las plaquetas maduras por citometría de flujo tras el marcaje fluorescente del contenido en RNA. Los colorante fluorescentes naranja de tiazol y auramina O, cuya fluorescencia aumenta después de la unión al RNA, han permitido el análisis por citometría de flujo del contenido en RNA de células anucleadas, incluyendo las plaquetas. Una caracterización más específica del proceso de trombopoyesis es posible basándose en el análisis del contenido relativo de RNA, relacionado con el grado de maduración de la población que contiene RNA. Además, la caracterización de la expresión de receptores en estas células puede permitir el análisis temprano del estado funcional de la maduración plaquetaria en la médula46.
El análisis de plaquetas reticuladas es de especial importancia en el diagnóstico de trombocitopenias y en la caracterización de la eficiencia de la terapia con trombopoyetina.
Detección de anticuerpos antiplaquetarios por citometría de flujo
La citometría de flujo puede ser utilizada en la identificación, cuantificación y caracterización de los anticuerpos unidos a la superficie de las plaquetas, proporcionando una información útil en el diagnóstico de disfunciones relativamente comunes, las trombocitopenias de naturaleza inmunitaria. La cuantificación por citometría de flujo de imunoglobulinas unidas a plaquetas usando antisueros IgG e IgM anti-humanos es un test muy sensible. Este análisis puede ser realizado directamente, para la detección de anticuerpos ya unidos a las plaquetas o, indirectamente, tras la incubación con plaquetas problema, para el "screening" de anticuerpos dirigidos contra la plaqueta.
Ventajas y limitaciones de la citometría de flujo en el estudio de las plaquetas
Ventajas
La citometría de flujo permite determinar rápidamente características específicas de un gran número de células individuales. En comparación con otras metodologías, la citometría de flujo tiene la ventaja de permitir analizar poblaciones plaquetarias en que las plaquetas son evaluadas una a una, posibilitando la identificación de subpoblaciones, mientras que los otros métodos reflejan la activación media de la población plaquetaria47.
Esta metodología permite evaluar tanto el estado de activación de las plaquetas circulantes como la reactividad plaquetaria. En la determinación de la activación plaquetaria in vivo, las dificultades resultantes de la posible activación ex vivo pueden ser evitadas por la fijación inmediata de las muestras de sangre, antes de proceder a su manipulación. Así pueden ser detectadas subpoblaciones parcialmente activadas menores del 1%.
Actualmente se dispone comercialmente de muchos anticuerpos y fluorocromos que permiten detectar numerosos parámetros plaquetarios, incluyendo diversos marcadores de activación. Los nuevos anticuerpos monoclonales desarrollados contra nuevos epítopos funcionales pueden ser incorporados fácilmente en los ensayos.
Los estudios de citometría requieren pequeñas cantidades de muestras, que pueden ser sangre entera, suspensiones de plaquetas lavadas, filtradas en gel o PRP.
Los estudios clínicos en que se utilizan suspensiones de plaquetas aisladas, tal como otros ensayos de la función plaquetaria, son potencialmente susceptibles de activación artefactual in vitro, resultante de los procedimientos de separación necesarios. En contraste, en el análisis de sangre entera la manipulación es mínima, evítando la activación artefactual y la posible pérdida de subpoblaciones plaquetarias.
Limitaciones
Los citómetros de flujo son aparatos muy caros, no estando aún disponibles en muchos laboratorios, principalmente en los de investigación básica.
La citometría de flujo no permite con facilidad una cuantificación absoluta de los antígenos detectados. Sin embargo, hay métodos actualmente disponibles para la cuantificación del número de anticuerpos unidos por célula.
Las plaquetas son muy pequeñas y los señales de FS y SS pueden no distinguirse bien del debris. La señal de fluorescencia también es más débil con respecto a marcajes fluorescente de otras células. Por esa razón la adquisición de los datos debe de ser efectuada con amplificación logarítmica. Sin embargo, los citómetros más recientes presentan resoluciones superiores, que permiten distinguir y estudiar con gran exactitud la población plaquetaria.
En ocasiones, hay dificultades en preservar el marcador, por ejemplo la P-selectina no se preserva bien, especialmente si es fijada.
La citometría de flujo solo mide la función de las plaquetas circulantes, mientras que los ensayos plasmáticos de β-TG y PF4 y los ensayos de metabolitos de TxA2 plasmáticos y urinarios también reflejan la activación plaquetaria en la pared del vaso y de las plaquetas recién separadas de circulación.
La elección de los parámetros adecuados para analizar el fenómeno de interés y la valoración de los resultados, teniendo en cuenta las condiciones experimentales subyacentes, serán posiblemente las principales limitaciones de la citometría de flujo en el estudio de la función plaquetaria.
Citometría de flujo en el estudio clínico de las plaquetas: Aplicaciones al Diagnóstico
La citometría de flujo proporciona una aproximación multiparamétrica para caracterizar las propiedades estructurales y funcionales de las plaquetas en condiciones normales. Esto permite establecer los rangos de normalidad, reconocer las plaquetas con comportamiento anormal y, en algunos casos, determinar el mecanismo de sus alteraciones. El número elevado de plaquetas analizado normalmente en una muestra permite la detección y cuantificación de subpoblaciones alteradas, que representan una pequeña fracción de la población total.
En los últimos años, la citometría de flujo empieza a ser aplicada como técnica multiparamétrica, para analizar el fenotipo y la función plaquetaria, incluyendo la activación. Sin embargo, los aspectos metodológicos y la elección de los parámetros biológicos adecuados están todavía en discusión.
Inmunofenotipaje de trombastenias
La citometría de flujo puede ser usada junto con anticuerpos monoclonales para cuantificar el número de complejos GPIIb/IIIa y GPIb/IX/V y así, evaluar la homozigosia o heterozigosia en la enfermedad de Bernard Soulier y en la Trombastenia de Glanzmann48. Permite, además, distinguir entre algunas variantes de trombastenia. Por ejemplo las plaquetas de indivíduos con la variante "cam" de la trombastenia contienen cantidades casi normales de GPIIb/IIIa. Sin embargo, se agregan deficientemente. Estas plaquetas no unen el anticuerpo PAC-1 y tampoco se unen al anticuerpo contra LIBS tras la adición de péptidos que contienen la secuencia Arg-Gly-Asp7.
Anomalías funcionales
Las disfunciones plaquetarias, incluyen las hereditarias y adquiridas. Estas anomalías son causas comunes de hemorragias, especialmente asociadas a traumatismos y cirugía.
La funcionalidad de las plaquetas se evalua tradicionalmente mediante ensayos de agregometría y adhesión. Sin embargo, estos ensayos solo indican si la función está inhibida. Por citometría de flujo pueden ser analizados varios parámetros bioquímicos de activación inducida por agonistas apropiados, proporcionando una información más especifica. Adicionalmente, la adhesion y agregación pueden ser seguidas por citometría de flujo midiendo la disminución de concentración de plaquetas individuales o la formación de agregados a través de cambios de las señales de FS y SS de partículas positivas para un marcador específico de la plaqueta (por ejemplo el CD41).
Las plaquetas de pacientes con trombastenia de Glanzmann presentan agregación reducida o ausente en respuesta a todos los agonistas, mientras que en el síndrome de Bernard Soulier las plaquetas presentan adhesión anormal. La citometría de flujo, además de constituir la técnica de elección para la detección de la expresión alterada o deficiente de glicoproteinas en la superficie plaquetaria permite investigar los defectos funcionales, consecuencia de las mismas.
La deficiencia hereditaria del acervo de almacenamiento de los gránulos densos, es una causa relativamente común de hemorragias moderadas, que no puede ser diagnosticada fiablemente por un ensayo normal de agregometría. El método convencional para establecer el diagnóstico es marcar las plaquetas con el colorante fluorescente mepacrina, especifico para los nucleotidos de adenina, y medir la fluorescencia de las plaquetas por microscopía. Este tipo de ensayo no son los ideales para la práctica clínica porque son tediosos, subjectivos y examinan un pequeño número de plaquetas. En contraste, esta deficiencia del almacenamiento de los gránulos densos puede ser detectada con precisión por un ensayo sencillo y rápido por citometría de flujo. Los resultados presentan una buena correlación con aquellos obtenidos por métodos de microscopía óptica de fluorescencia, pero la detección es mejorada porque la fluorescencia se determina en un gran número de plaquetas. Una deficiencia adquirida de los gránulos densos ocurre a consecuencia de disfunciones mieloproliferativas y en la fase final de insuficiencia renal.
El síndrome de plaquetas grises, caracterizado por trombocitopenia y una marcada disminución o ausencia de gránulos a normales, puede ser diagnosticada por una baja expresión de P-selectina o trombospondina en la superficie plaquetaria, tras la estimulación con agonistas fuertes.
La citometría de flujo también puede ser informativa en el estudio del síndrome de Scott, una deficiencia en la actividad procoagulante de la plaqueta. Las plaquetas de estos individuos son deficientes en la formación de micropartículas y en la expresión de lugares de unión para el factor Va, tanto en las micropartículas, como en las plaquetas. En contrapartida, la activación de GP IIb/IIIa y la expresión de P-selectina son normales7.
Las plaquetas hiperreactivas y/ o activadas in vivo, asociadas a la hipercoagulabilidad y trombosis son también una categoría de plaquetas con función anormal a la cual se hará referencia más adelante.
Aloanticuerpos y autoanticuerpos
Las trombocitopenias (disminución de la cifra de plaquetas sanguíneas por debajo de 150x109/L) pueden ser producidas por diversos factores etiológicos. Entre ellos, los más frecuentes son los de naturaleza inmunitaria. En enfermedades como la trombocitopenia autoinmune (TAI), la trombocitopenia neonatal aloimune (TNA), púrpura trombocitopenia postransfusional (PTP), refractariedad a las transfusiones de plaquetas y en la trombocitopenia inducida por fármacos, los anticuerpos anti-plaquetarios son los principales responsables de la eliminación acelerada de plaquetas49.
Los aloanticuerpos se dirigen contra determinantes antigenicos alotípicos que están presentes en la membrana plaquetaria de algunos individuos y ausentes en la de otros. Estos incluyen los anticuerpos para los aloantígenos clasificados en el sistema HPA (Human Platelet Antigen) y anticuerpos contra moléculas del sistema HLA. Por ejemplo el antígeno HPA-1 se encuentra implicado en la mayoría de las TNA y PTP49.
Los autoanticuerpos reconocen determinantes antigénicos que se expresan en las plaquetas de todos aquellos individuos que no presentan anomalías en las GPs de la membrana plaquetaria La mayoría de los anticuerpos en pacientes con púrpura trombocitopénica autoinmune reconocen uno o más epítopos localizados en GPIIb/IIIa. Ocasionalmente, estos anticuerpos reconocen epítopos en GPIb, GPIa, etc.49. Uno de los casos mejor conocidos es la trombocitopenia inducida por heparina, la cual es causada por la presencia de plaquetas activadas después del tratamiento con heparina. El PF4 en la superficie de las plaquetas activadas se une a la heparina, formando así un complejo antigénico.
La citometría de flujo puede ser utilizada en la identificación y cuantificación de las imunoglobulinas unidas a la membrana plaquetaria y, adicionalmente, en la detección de la activación plaquetaria inducida por estos anticuerpos. Esta metodología proporciona así, una información substancial en el diagnóstico de las trombocitopenias inmunes, con varias ventajas, sobre todo la posibilidad de estudiar casos de trombocitopenia severa con volúmenes de sangre relativamente pequeños.
Citometría de flujo en el estudio clínico de las plaquetas: Aplicaciones al Pronóstico
Las alteraciones en la activación y agregación plaquetaria pueden conducir tanto a situaciones protrombóticas como a estados hemorrágicos, por lo que se consideran importantes factores de riesgo de diversas enfermedades cardiovasculares. Por esa razón el estudio de marcadores bioquímicos tempranos de activación puede ser relevante para evaluar el estado de activación de las plaquetas circulantes (activación in vivo) y la reactividad alterada, así como para proporcionar factores de riesgo predictivos en las disfunciones hemostáticas y sus terapias.
Una vez establecido el diagnóstico celular, el pronóstico del proceso patológico puede evaluarse sobre la base de un ensayo citométrico predictivo, normalmente en coordinación con parámetros clínicos bien establecidos.
A) Detección de plaquetas activadas circulantes
La activación plaquetaria, con la consecuente formación de trombos blancos, desempeña un papel importante en la patogénesis de la enfermedad coronaria arterial, incluyendo la angina inestable. Por otra parte la activación plaquetaria en respuesta a una lesión local o alteraciones hemodinámicas agudas se considera el evento que precipita la oclusion final del vaso, llevando al infarto de miocardio50-51.
Se ha descrito la asociación entre activación plaquetaria in vivo y/o hiperreactividad con la angina inestable, infarto de miocardio y otras situaciones clínicas comunes incluyendo angioplastia, by-pass cardiopulmonar, ataques isquémicos transitorios, diabetes mellitus, hiperlipoproteinemia, stress emocional y ejercicio intenso52. En el caso de la diabetes, el fenotipo activado refleja un defecto metabólico intrínseco.
Considerando la citometría como un ensayo predictivo, se ha observado que el grado de activación plaquetaria in vivo, evaluado a través de la expresión de diversos marcadores de activación, está directamente relacionado con el proceso trombótico. La mayoría de los estudios de la activación plaquetaria mediante citometría de flujo consisten en el análisis inmunofenotípico de las glicoproteínas de superficie que se expresan después de la activación de las plaquetas con estímulos específicos, como la P-selectina (CD62P), CD63, trombospondina, GPIIbIIIa activada53,54. En ausencia de la adición de agonistas exógenos se ha detectado activación de plaquetas circulantes, usando anticuerpos específicos para las referidas glicoproteínas, sobre todo para CD62. Para prevenir la activación artefactual de las plaquetas in vitro, durante la obtención y almacenamiento, las muestras de sangre son estabilizadas con antagonistas de la activación plaquetaria y se fijan, en el caso que no sea posible realizar el análisis en un corto espacio de tiempo.
El límite mínimo de detección de activación plaquetaria por citometría de flujo, utilizando anticuerpos para GPIIbIIIa activada o CD62P, se considera aproximadamente el 1% de plaquetas activadas en muestras estimuladas y no estimuladas6. Sin embargo, en diversas situaciones clínicas con activación plaquetaria in vivo, la subpoblación de plaquetas activadas puede ser menor y, en estos casos, el ensayo no es suficientemente sensible.
Estudios recientes han demostrado que CD62P puede perderse rápidamente in vivo después de la desgranulación de la plaqueta52, limitando así significativamente su aplicación predictiva a los estudios ex vivo e in vitro. Sin embargo, la expresión transitoria no es única para CD62P. En particular, la unión del fibrinógeno se revierte aún mas rápidamente cuando las plaquetas son estimuladas in vitro, el aumento de la expresión de GPIIbIIIa también es reversible y, probablemente, también lo son todos los neoepitopos asociados a la activación de GPIIbIIIa. El período de tiempo que estos marcadores permanecen expresados en la plaqueta in vivo no ha sido todavía determinado. Además, la cuantificación precisa de las glicoproteínas plaquetarias se dificulta con la muy variable acesibilidad de los epitopos tras la redistribución del sistema canalicular abierto inducida por la activación de la plaqueta.
La sensibilidad y especificidad de los ensayos de citometría de flujo, por consiguiente, deberá ser determinada a través de su correlación con parámetros clínicos de enfermedad en un gran número de pacientes.
B) Activabilidad ex vivo
La hiperreactividad plaquetaria puede ser monitorizada por diversos ensayos que miden aspectos distintos de la activación plaquetaria. Muchos estudios han mostrado que la reactividad plaquetaria está aumentada en pacientes con varios tipos de disfunciones arteriales trombóticas55. Los mecanismos subyacentes a la hiperreactividad plaquetaria en estos pacientes son inciertos. La hiperreactividad puede ser debida a una excesiva producción de agonistas, receptores hiperreactivos, o anomalías en los mecanismos de transducción de la señal. Una respuesta incrementada tras estimulación se ha considerado la manifestación de un bajo grado de preactivación plaquetaria en los sindromes pretrombóticos.
Estas anomalías pueden ser investigadas convenientemente por citometría de flujo, estimulando las plaquetas con cantidades definidas de agonistas. La utilización de agonistas totalmente sintéticos y, por eso, altamente controlables, como el ADP, TRAPs o los análogos estables del tromboxano A2, permiten la activación in vitro con elevado grado de reproducibilidad. Estos ensayos pueden ser realizados en sangre entera diluída en condiciones no agregantes, posibilitando el análisis de grados altos de activación sin la pérdida de subpoblaciones6. Incluso la activación inducida por la trombina, uno de los activadores fisiológicos mas importantes, puede ser determinada directamente en sangre entera usando el tetrapéptido sintético glicil-L-prolil-L-arginil-L-prolina (GPRP). El GPRP inhibe la polimerización de fibrina y la agregación plaqueta-plaqueta55, permitiendo el análisis de plaquetas individuales. Un aspecto interesante a considerar es que los cambios en los marcadores pueden tener diferentes significados. Por ejemplo, el antígeno CD63 requiere estimulaciones muy fuertes para expresarse, CD62P se expresa con estimulaciones moderadas y la expresión del epítopo reconocido por el PAC-1, ocurre con estimulaciones débiles.
Aplicando la citometría de flujo es posible evaluar simultáneamente la agregación y la secreción. Ault et al.57 describieron un método que permitió estudiar la relación entre la secreción y la agregación plaquetaria, en sangre entera, por citometría de flujo. Un anticuerpo monoclonal específico para CD62P, marcado con ficoeritrina fue usado para evaluar la secreción, y un anticuerpo específico para GPIb, marcado con fluoresceína-isotiocianato (FITC), fue usado para distinguir las plaquetas de los agregados plaquetarios.
La inestabilidad de las plaquetas limita significativamente los estudios de reactividad plaquetaria in vitro, ya que deben ser efectuados en un corto espacio de tiempo después de la punción venosa.
C) Monitorización de la terapeutica
Con la tendencia actual a usar agentes antiplaquetarios como profilaxis para los eventos trombóticos, son necesarias metodologías para evaluar la eficacia de la terapia. Para monitorizar los efectos de un agente anti-plaquetario, se determinan los efectos en las respuestas plaquetarias inducidas por los agonistas. Estos ensayos deben ser sensibles a los mecanismos de actuación del fármaco, siendo la especificidad y sensibilidad conseguidas lo que determina la elección de los agonistas apropiados. Por ejemplo, estimulando las plaquetas con ácido araquidónico o bajas concentraciones de colágeno, la agregación plaquetaria es principalmente dependiente de la formación de TXA2, y estas serán las condiciones ideales para probar el efecto de la aspirina y otros fármacos antiinflamatorios no esteroides. Utilizando el ADP como estímulo, se puede determinar el efecto inhibidor de la ticlopidina en las respuestas plaquetarias58.
Recientemente ha surgido una nueva clase de agentes antitromboticos, los antagonistas de GPIIbIIIa. El bloqueo del receptor GPIIbIIIa impide la unión del fibrinógeno y, consecuentemente, inhibe la agregación plaquetaria, la fase final de la formación del trombo. Se han desarrollado una gran variedad de anticuerpos, péptidos y otros tipos de moléculas que bloquean el receptor GPIIbIIIa y han sido incluidos en ensayos clínicos59. Igualmente, se han desarrollado métodos de análisis por citometría de flujo para monitorizar estos tratamientos, de particular importancia porque estos fármacos poseen un rango terapeutico muy estrecho. El análisis de la unión de anticuerpos60, de fibrinógeno61 o de la expresión de LIBS13 puede usarse para cuantificar la inhibición inducida por estos agentes.
La medida de los marcadores bioquímicos por citometría de flujo puede mostrar cambios más tempranos en el estado de activación de las plaquetas que los detectados por ensayos fisiológicos. Estos ensayos pueden detectar alteraciones en la bioquímica plaquetaria en respuesta al fármaco, aunque no haya alteraciones en la agregación. La citometría de flujo, además de monitorizar el efecto global del tratamiento, permite también detectar subpoblaciones heterogéneas en la respuesta a la terapeutica. La capacidad multiparamétrica de la citometría de flujo proporciona una excelente metodología para investigar la utilidad de estos ensayos en la elección de las dosis antitrombóticas óptimas de fármaco, para monitorizar la eficacia y la seguridad de la terapeutica. Por otra parte, la identificación de individuos sensibles o resistentes a una terapia determinada, puede permitir que los individuos resistentes se beneficien de un tratamiento preventivo con otro agente antiplaquetario.
D) Control de calidad de plaquetas almacenadas:
Varios estudios por citometría de flujo en los que se determinó la expresión de CD62P, CD63 y GPIIbIIIa en plaquetas almacenados en el banco antes de la transfusión, revelaron que se produce activación plaquetaria dependiente del tiempo62,63. Estas alteraciones están correlacionadas con cambios en la morfología plaquetaria y liberación de b-TG62. Se sugirió que la presencia de CD62P en la superficie de la plaqueta puede ser útil como una medida de control de calidad, basado en correlaciones entre la expresión basal de CD62P en la superficie plaquetaria y en las plaquetas almacenadas en el banco y, por otra parte, con la cifra de plaquetas posttransfusión, la sobrevida plaquetaria y el aclaramiento del receptor de las plaquetas CD62P positivas62.
Las plaquetas almacenadas presentan defectos funcionales, como se demostró, por citometría de flujo, puesto que durante el almacenamiento disminuye la movilización de Ca2+ citosólico inducida por la trombina, y se forma una subpoblación completamente insensible30.
Bibliografía
1. Metzelaar MJ, Sixma JJ, Nieuwenhuis HK. Detection of platelet activation using activation specific monoclonal antibodies. Blood Cells 1990; 16: 85-96. [ Links ]
2. Newman PJ, Allen RW, Kahn RA, Kunicki TJ. Quantification of membrane glycoprotein IIIa on intact human platelets using the monoclonal antibody AP-3. Blood 1985; 65 (1): 227-232. [ Links ]
3. Adelman B, Michelson AD, Handin RI, Ault KA. Evaluation of platelet glycoprotein Ib by fluorescence flow cytometry. Blood 1985; 66 (2): 423-427. [ Links ]
4. Michelson AD. Thrombin-induced down-regulation of the platelet membrane glycoprotein Ib-IX complex. Sem Thromb Haemost 1992; 18 (1): 18-27. [ Links ]
5. McEver RP, Martin MN. A monoclonal antibody to a membrane glycoprotein binds only to activated platelets. J Biol Chem 1984; 259 (15): 9799-9804. [ Links ]
6. Shattil SJ, Cunningham M, Hoxie JA. Detection of activated platelets in whole blood using activation-dependent monoclonal antibodies and flow cytometry. Blood 1987; 70 (1): 307-315. [ Links ]
7. Abrams C, Shattil SJ. Immunological detection of activated platelets in clinical disorders. Thromb Haemost 1991; 65 (5): 467-473. [ Links ]
8. Sims PJ, Ginsberg MH, Plow EF, Shattil SJ. Effect of platelet activation on the conformation of the plasma membrane glycoprotein IIb/III complex. J Biol Chem 1991; 266: 7345-7352. [ Links ]
9. Frojmovic MM. Flow Cytometric analysis of platelet activation and fibrinogen binding. Platelets 1996; 7: 9-21. [ Links ]
10. Ugarova TP, Budzynski AZ, Ruggeri ZM, Ginsberg MH, Plow EF. Conformational changes in fibrinogen elicited by its interaction with platelet membrane glycoprotein GPIIb/IIIa. J Biol Chem 1993; 268: 21080-21087. [ Links ]
11. Ginsberg MH, Frelinger AL, Lam SC-T, Forsyth J, McMillan R, Plow EF Shattil SJ. Analysis of platelet aggregation disorders based on flow cytometric analysis of membrane glycoprotein IIb-IIIa with conformation-specific monoclonal antibodies. Blood 1990; 76 (10): 2017-2023. [ Links ]
12. Mandoro TH, Walt CD, White MM, Jennings LK. Selective induction of a GPIIb/IIIa LIBS by fibrinogen and vWF. Blood 1996; 88: 3824-3830. [ Links ]
13. Jennings LK, White MM. Expression of ligand-induced binding sites on glycoprotein IIb/IIIa complexes and the effect of various inhibitors. Am Heart J 1998; S179-S183. [ Links ]
14. Corash L. Measurement of platelet activation by fluorescence-activated flow cytometry. Blood Cells 1990; 160: 97-108. [ Links ]
15. Stenberg PA, McEver R, Shuman MA, Jaques YV, Baiton DF. A platelet alpha-granule membrane protein (GMP-140) isexpressed on the plasma membrane after activation. J Cell Biol 1985; 101: 880-886. [ Links ]
16. Wu G, Xi X, Li P, Chu X, Ruan C. Preparation of a monoclonal antibody, SZ-51, that recognizes an a-granule membrane protein (GMP-140) on the surface of activated human platelets. Nouv Rev Fr Hematol 1990; 32: 231-235. [ Links ]
17. Parmentier S, McGregor L, Catimel B, Leung LLK, McGregor JL. Inhibition of platelet functions by a monoclonal antibody (LYP20) directed against a granule membrane glycoprotein (GMP140/PADGEM). Blood 1991; 77 (8): 1734-1739. [ Links ]
18. Metzelaar MJ, Heijnen HFG, Sixma JJ, Nieuwenhuis HK. Identification of a 33-Kd protein associated with the a-granule membrane (GMP-33) that is expressed on the surface of activated platelets. Blood 1992; 79 (2): 372-379. [ Links ]
19. Nieuwenhuis HK, Oosterhout JJG, Rozemuller E, Iwaarden F, Sixma JJ. Studies with a monoclonal antibody aginst activated platelets: evidence that a secreted 53.000-molecular weight lysosome-like granule protein is exposed on the surface of activated platelets in the circulation. Blood 1987; 70 (3): 838-845. [ Links ]
20. Capitanio AM, Niewiarowski S, Rucinski B, Tuszynsky GP, Cierniewski CS, Hershock D, Kornecki E. Interaction of platelet factor 4 with human platelets. Biochem Biophys Acta 1985; 839: 161-173. [ Links ]
21. Rubinstein E, Urso I, Boucheix C, Carroll RC. Mechanisms for expression of thrombospondin on the platelet surface. Semin Thromb Haemost 1987; 13: 307 -316. [ Links ]
22. Abrams CS, Ellison N, Budzynski AZ, Shattil SJ. Direct detection of activated platelets and platelet-derived microparticles in humans. Blood 1990; 75 (1): 128-138. [ Links ]
23. Legrand C, Dubernard V, Rabhi-Sabile S, Morandi da Silva V. Functional and clinical significance of thrombospondin. Platelets 1997; 8: 211-223. [ Links ]
24. Sims PJ, Faioni EM, Wiedmer T, Shattil SJ. Complement proteins C5b-9 cause release of membrane vesicles from the platelet suface that are enriched in the membrane eceptor for coagulation factor Va and express prothrombinase activity. J Biol Chem 1988; 263 (34) 18205-18212. [ Links ]
25. Dachary-Prigent J, Pasquet J-M, Freyssinet J-M, Nurden AT. Calcium involvement in aminophosphoslipid exposure and microparticle formation during platelet activation: a study using Ca2+- ATPase inhibitors. Biochemistry 1995; 34: 11625-11634. [ Links ]
26. Dachary-Prigent J, Toti F, Satta N, Pasquet J-M, Uzan A, Freyssinet J-M. Physiopathological significance of catalytic phospholipids in the generation of thrombin. Semin Thromb Haemost 1996; 22 (2): 157-163. [ Links ]
27. Salzman EW, Ware JA. Ionized calcium as an intracellular messenger in blood platelets. Prog Hemost Thromb 1989; 9: 177-202. [ Links ]
28. Jennings LK, Dockter ME, Wall CD, Fox CF, Kennedy DM. Calcium Mobilization in human platelets using indo-1and flow cytometry. Blood 1989; 74 (8): 2674-2680. [ Links ]
29. Davies TA, Weil GJ, Simons ER. Simultaneous flow cytometric measurements of thrombin-induced cytosolic ph and Ca2+ fluxes in human platelets. J Biol Chem 1990; 265 (20): 11522-11526. [ Links ]
30. Fijnheer R, Homburg CHE, Hooibrink B, Boomgaard MN, Korte D, Roos D. Loss of thrombin-induced Ca2+ mobilization in subpopulation of platelets during storage. Thromb Haemost 1991; 66 (3): 350-354. [ Links ]
31. Kyle PM, Jackson MC, Buckley DC, Swiet M, Redman CWG. Platelet intracellular free calcium response to aeginine vasopression is similar in preeclampsia and normal pregnancy. Am J Obstet Gynecol 1995; 172 (2): 654-660. [ Links ]
32. Monteiro MC, Sansonetty F, Gonçalves MJ, O'Connor JE. Flow cytometric kinetic assay of calcium mobilization in whole blood platelets using Fluo-3 and CD41. Cytometry 1999; 35: 302-310. [ Links ]
33. Tschoepe D, Spangenberg P, Esser J, Schwippert B, Kehrel B, Roesen P, Gries FA. Flow-cytometric detection of surface membrane alterations and concomitant changes in the cytoskeletal actin status of activated platelets. Cytometry 1990; 11: 652-656. [ Links ]
34. Chow S, Hedley D. Flow cytometric measurement of intracellular pH. Current Protocols in Cytometry 1997; 9.3.1-9.3.10. [ Links ]
35. Atsumi T, Sugita K, Kohono M, Takahashi T, Ueha T. Simultaneous measurement of Ca2+ and pH by laser cytometry using fluo-3 and SNARF-1. Cytometry 1996; 24: 99-105. [ Links ]
36. Cavallini L, Coassin M, Alexandre A. Diacyglycerol mediates the thrombin-induced, protein kinase C and Ca2+ independent activation of the Na+/H+ exchanger in platelets. FEBS 1997; 400: 99-102. [ Links ]
37. Nofer J-R, Tepel M, Kehrel B, Walter M, Seedorf U, Assmann G, Zidek W. High density lipoproteins enhance the Na+/H+ antiport in human platelets. Thromb Haemost 1996; 75 (4): 635-641. [ Links ]
38. Nofer J-R, Tepel M, Kehrel B, Wierwille S, Walter M, Seedorf U, Zidek W, Assmann G. Low-density lipoproteins inhibit the Na+/H+ antiport in human platelets. Circulation 1997; 95 (6): 1370-1377. [ Links ]
39. Oda A, Daley J, Cabral C, Kang J, Smith M, Salzman EW. Heterogeneity in filamentous actin content among individual human blood platelets. Blood 1992; 79 (4): 920-927. [ Links ]
40. Gordon N, Thom J, Cole C, Baker R. Rapid detection of hereditary and acquired platelet storage pool deficiency by flow cytometry. Brit J Haematol 1995; 89: 117-123. [ Links ]
41. Horne WC, Simons ER. Probes of transmembrane potential in platelets: changes in cyanine dye fluorescence in response to aggregation stimuli. Blood 1978; 51: 741-749. [ Links ]
42. Greenberg-Sepersky SM, Simons ER, White JG. Studies of platelets from patients with grey platelet syndrome. Br J Haematol 1985; 59 (4): 603-609. [ Links ]
43. Iuliano L, Colavita AR, Leo R, Praticò D, Violo F. Oxygen free radicals and platelet activation. Free Radical Biol Med 1997; 22 (6): 999-1006. [ Links ]
44. Leo R, Praticò D, Iuliano L, Pulcinelli FM, Ghiselli A, Pignatelli P, Colavita AR, FitzGerald GA, Violi F. Platelet activation by superoxide anion and hydroxyl radicals intrinsically generated by platelets that had undergone anoxia and then reoxygenated. Circulation 1997; 95 (4): 885-891. [ Links ]
45. Leoncini G, Signorello MG, Piana A, Carrubba M, Armani U. Hydrogen peroxide formation in platelets of patientes with non-insulin-dependent diabetes mellitus. Platelets 1998; 9: 213-217. [ Links ]
46. Matic GB, Rothe G, Schmitz G. Flow cytometric analysis of reticulated platelets. Current Protocols in Cytometry 1998; 7.10.1-7.10.6. [ Links ]
47. Muirhead KA, Horan PK, Post G. Flow cytometry: present and future. Biotechnology 1985; 3: 337-356. [ Links ]
48. Jennings LK, Ashmun RA, Wang WC, Dockter ME. Analysis of human platelet glycoproteins IIb-IIIa and glanzmann's trombasthenia in whole blood by cytometry. Blood 1986; 68 (1): 173-179. [ Links ]
49. Pujol-Moix N. In: Trombocitopenia. Mosby, Madrid, 1995; pp 32-40. [ Links ]
50. Fitzgerald DJ, Roy L, Catella F, Gerald GAF. Platelet activation in unstable coronary disease. New Engl J Med 1986; 315 (16): 983-989. [ Links ]
51. Flores NA, Sheridan DJ. The pathophysiological role of platelets during myocardial ischaemia. Cardiovasc Res 1994; 28 (3): 295-302. [ Links ]
52. Michelson AD: Flow cytometry: a clinical test of platelet function. Blood 1996; 87: 4925-4936. [ Links ]
53. Goodall AH. Platelet activation during preparation and storage of concentrates: detection by flow cytometry. Blood Coag Fibrinolysis 1991; 2: 377-382. [ Links ]
54. Metzelaar MJ, Clevers HC. Lysossomal membrane glycoproteins in platelet. Thromb Haemost 1992; 68 (4): 378-382. [ Links ]
55. Grand R. Platelet activation and arterial thrombosis. Lancet 1994; 344: 991-994. [ Links ]
56. Michelson A, Ellis PA, Barnard MR, Matic GB, Viles AF, Kestin AS. Down-regulation of the platelet surface glycoprotein Ib-IX complex in whole blood stimulated by thrombin, adenosine diphosphate, or an in vivo wound. Blood 1991; 77 (4): 770-779. [ Links ]
57. Ault KA, Rinder HM, Mitchell JG, Rinder CS, Lambrew CT, Hillman RS. Correlated measurement of platelet release and aggregation in whole blood. Cytometry 1989; 10: 448-455. [ Links ]
58. Schmitz G, Rothe G, Ruf A, Barlage S, Tschope D, Clemetson KJ, Goodall AH, Michelson AD, Nurden AT, Shankey TV. European working group on clinical cell analysis: Consensus protocol for the flow cytometric characterisation of platelet function. Thromb Haemost 1998; 79: 885-896. [ Links ]
59. Fergunson JJ, Lau TK. New antiplatelet agents for acute coronary syndromes. Am Heart J 1998; 135 (5). S194-S199. [ Links ]
60. Konstantopoulos K, Grotta JC, Sills C, Wu KK. Shear-induced platelet agregation in normal subjects and stroke patients. Thromb Haemost 1995; 74 (5): 1329-1334. [ Links ]
61. Cox D. Methods for monitoring platelet function. Am Heart J 1998; S160-S169. [ Links ]
62. Fijnheer R, Modderman PW, Veldman H, Ouwehand WH, Nieuwenhuis HK, Roos D, De Kort D. Detection of platelet activation with monoclonal antibodies an flow cytometry. Changes during platelet storage. Transfusion 1990; 30 (1): 20-25. [ Links ]
63. Matsubayashi H, Weidner J, Miraglia CC, McIntyre JÁ. Platelet membrane early activation markers during prolonged storage. Thromb Res 1999; 151-160. [ Links ]

