










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkArchivos de la Sociedad Española de Oftalmología
versión impresa ISSN 0365-6691
Arch Soc Esp Oftalmol vol.85 no.12 dic. 2010
Expresión de marcadores embrionarios en células mesenquimales de pterygium
Expression of embryonic markers in pterygium derived mesenchymal cells
G. Pascuala,d, M.A. Montesb, C. Pérez-Ricob,c, B. Pérez-Kohlera, J.M. Bellónc,d y J. Bujána,d
aDepartamento de Especialidades Médicas, Universidad de Alcalá de Henares, Alcalá de Henares, Madrid, España
bServicio de Oftalmología, Hospital Universitario Príncipe de Asturias, Alcalá de Henares, Madrid, España
cDepartamento de Cirugía, Universidad de Alcalá de Henares, Alcalá de Henares, Madrid, España
dCentro de Investigación en Red en Bioingeniería, Biomateriales y Nanomedicina (CIBER-BBN), Madrid, España
Dirección para correspondencia
RESUMEN
Propósito: La destrucción de la barrera del epitelio limbal es el mecanismo más importante en la formación del pterygium. Se piensa que proviene de la activación y proliferación de células madre (stem cells) del epitelio limbal. El objetivo de este estudio ha sido determinar la presencia de células mesenquimáticas indiferenciadas (stem cells) en cultivos celulares obtenidos de pterygium humano.
Métodos: Las células de 6 pterygium humanos fueron aisladas mediante explantos y cultivadas en amniomax. Al alcanzar la confluencia, las células fueron sembradas en microplacas de 24 pocillos. Los cultivos celulares fueron estudiados en el 2.o subcultivo. Se analizó mediante inmunofluorescencia la expresión de los marcadores de células madre embrionarias, OCT3/4 y CD9. Los diferentes fenotipos celulares fueron caracterizados con los anticuerpos monoclonales anti-CD31, α-actina y vimentina.
Resultados: Toda la población celular obtenida de pterygium mostró expresión para vimentina. Menos de un 1% de las células presentaron expresión para los marcadores CD31 y α-actina. La mayor parte de la población celular fue positiva para los marcadores embrionarios OCT3/4 y CD9.
Conclusión: La población celular expandida a partir de pterygium está compuesta por células de fenotipo mesenquimal, que muestran expresión de marcadores embrionarios OCT3/4 y CD9. Esta población, con un alto grado de indiferenciación, podría ser responsable de la alta tasa de recurrencia del pterygium tras su extirpación quirúrgica.
Palabras clave: Pterygium. Células madre. Marcadores embrionarios.
ABSTRACT
Background: Destruction of the limbal epithelium barrier is the most important mechanism of pterygium formation (conjunctiva proliferation, encroaching onto the cornea). It is thought to arise from activated and proliferating limbal epithelial stem cells. The objective of this study is to evaluate the presence of undifferentiated mesenchymal cells (stem cells) in cultured cells extracted from human pterygium.
Material and methods: Cells from 6 human pterygium were isolated by explantation and placed in cultures with amniomax medium. Once the monolayer was reached the cells were seeded onto 24 well microplates. The cells were studied in the second sub-culture. The immunohistochemical expression of different embryonic stem cell markers, OCT3/4 and CD9, was analysed. The differentiated phenotypes were characterised with the monoclonal antibodies anti-CD31, α-actin and vimentin.
Results: All the cell populations obtained from pterygium showed vimentin expression. Less than 1% of the cells were positive for CD31 and α-actin markers. The majority of the cell population was positive for OCT3/4 and CD9.
Conclusions: The cell population obtained from pterygium expressed mesenchymal cell phenotype and embryonic markers, such us OCT3/4 and CD9. This undifferentiated population could be involved in the large recurrence rate of this type of tissue after surgery.
Key words: Pterygium. Stem cells. Embryonic markers.
Introducción
El pterygium se caracteriza por un crecimiento fibrovascular de la conjuntiva que se extiende a través del limbo, invadiendo la córnea1. Se trata de una afección de la superficie ocular relativamente frecuente, más prevalente en sujetos de países con climas cálidos, donde la exposición a la radiación ultravioleta es más intensa2. Histopatológicamente se trata de un epitelio conjuntival atrófico, bajo el cual existe un tejido engrosado, hipertrófico, de tipo conectivo con degeneración elastoide3. La extensión y severidad de dicho tejido fibrovascular se consideran factores predictivos de recurrencia tras la cirugía.
La etiopatogenia del pterygium no es del todo conocida, aunque se ha relacionado con factores ambientales como la radiación ultravioleta, irritación e inflamación crónica. En la actualidad se acepta que el desarrollo del pterygium tiene lugar en dos fases, la rotura inicial de la barrera epitelial córneo-conjuntival y el crecimiento del pterygium. La radiación ultravioleta dañaría las células madre limbares, de forma directa por su efecto fototóxico y de forma indirecta por la formación de radicales libres que son altamente lesivos4. En estas células madre dañadas existe una expresión alterada del gen de supresión tumoral p53, que causaría la modificación de las vías de apoptosis5 y además, estas células limbares degeneradas son capaces de incrementar la expresión de las metaloproteinasas de la matriz (MMP-1, MMP-2 y MMP-9) que serían responsables de aumentar la rotura de la barrera epitelial córneo-conjuntival6. En el crecimiento del pterygium, han sido implicados diversos factores de crecimiento fibroangiogénico, como el factor de crecimiento fibroblástico (FGF), factor de crecimiento epidérmico ligado a la heparina (HB-EGF), factor de crecimiento tipo insulina ligado a proteína 2 (IGFBP), factor de crecimiento epidérmico (EGF), factor de crecimiento del tejido conectivo (CTGF) y sobre todo el factor de crecimiento endotelial vascular (VEGF)7-10.
El objetivo de este estudio ha sido determinar la presencia de células mesenquimales indiferenciadas en cultivos celulares obtenidos de pterygium humano, con idea de mejorar los conocimientos de los mecanismos fisiopatológicos implicados en este proceso degenerativo.
Material y métodos
Selección de la muestra
Un total de 6 muestras procedentes de 6 pacientes con pterygium fueron incluidas en el estudio. El consentimiento informado fue firmado por todos los participantes. Los criterios de inclusión fueron los siguientes: 1) pterygium primario carnoso y pterygium activo que invadía en más de 3mm la córnea; 2) no cirugías ni traumatismos previos en el ojo afectado, y 3) no padecer enfermedades oculares y sistémicas que pudieran alterar la morfología de la superficie ocular.
Intervención quirúrgica
En todos los casos se realizó una resección simple del pterygium con aplicación intraoperatoria de mitomicina C. La anestesia conjuntival se indujo con la instilación de colirio de clorhidrato de tetracaína (Colirio anestésico doble®, Alconcusí, Barcelona, España) combinada con una inyección subconjuntival de 0,5ml de lidocaína al 2% con epinefrina al 1:200.000 en el cuerpo del pterygium, usando una aguja de 25-G. La cabeza del pterygium fue disecada de la córnea con un bisturí Bard-Parker n.o 15 y el cuerpo del pterygium junto con la tenon subyacente, fue escindido con tijeras de Wescott, dejando la esclera expuesta. La mitomicina C 0,02% (0,2mg/ml) fue aplicada directamente sobre el lecho escleral durante 1 minuto, a continuación la superficie ocular fue irrigada con solución salina balanceada. Las suturas conjuntivales (7-0 vicryl, Ethicon) fueron extraídas a los 7 días de la cirugía. Se prescribió tobramicina y dexametasona tópica (Tobradex®, Alconcusí, Barcelona, España), 4 veces al día durante 1 mes. El tejido extraído fue inmediatamente sumergido en medio mínimo esencial (MEM) estéril y almacenado a 4oC hasta su traslado al laboratorio.
Obtención de la población celular
Bajo condiciones de esterilidad, utilizando una cabina de flujo laminar de tipo II (Telstar AV 30/70, Telstar SA, Madrid, España), el tejido extraído quirúrgicamente fue cortado, con ayuda de un bisturí, en piezas de 1mm2. Estos explantos fueron depositados sobre el fondo de frascos de cultivo tipo Roux de 25cm2 de superficie (Nuclon-Intermed, Nunc A/S, Roskildo, Denmark), a los que se les añadió 3ml de medio de cultivo amniomax (Gibco BRL). Los frascos de cultivo fueron incubados a 37oC en presencia del 5% de CO2 en una estufa de cultivo celular. El medio de cultivo fue reemplazado 2 veces por semana. Una vez que las células crecieron hasta la confluencia, fueron subcultivadas mediante tratamiento enzimático con tripsina-EDTA (Gibco BRL). Las células obtenidas del 1er subcultivo fueron utilizadas en los experimentos siguientes.
Caracterización de la población celular
Las células procedentes del primer subcultivo fueron sembradas sobre cubreobjetos estériles en microplacas de 24 pocillos. Por un lado fueron fijadas durante 30 minutos en líquido de Bouin, aclaradas durante 10 minutos en etanol 70o y lavadas con tampón fosfato salino (PBS). Las células procedentes de pterygium fueron sometidas a ensayos de inmunofluorescencia con anticuerpos primarios anti-α-actina de músculo liso, anti-vimentina y anti-CD31 (Sigma, St. Louis, Mo., USA). Se empleó un anticuerpo secundario conjugado con FITC (Sigma, St. Louis, Mo., USA). Los núcleos fueron contrastados con 4´,6-diamino-2-phenylindole-dihydrochloride (DAPI).
La identificación de marcadores característicos de células madre embrionarias fue realizada en células fijadas con paraformaldehído 4% durante 20 minutos a 4oC. Los anticuerpos empleados fueron: anti-OCT3/4 y anti-CD9 (R&D System, PAIS). Se utilizaron anticuerpos secundarios conjugados con FITC y rodamina (Jackson Laboratories Inc. West Grove, USA). En todos los casos, los núcleos fueron contrastados con 4´,6-diamino-2-phenylindole-dihydrochloride (DAPI). Los controles negativos se realizaron sustituyendo el anticuerpo primario por una solución de PBS. Los cultivos celulares obtenidos de pterygium fueron observados y realizado el recuento en un equipo de microscopia confocal Leica SP5 (Leica, Germany).
Resultados
En todos los casos se pudo observar que a los 2 días de cultivo empezaban a aparecer células que partían de los diferentes explantos de pterygium depositados sobre el frasco de cultivo y éstas incrementaban la población rápidamente (Figura 1). El 100% de la población celular obtenida del pterygium presentó positividad para la proteína vimentina. Los filamentos de vimentina aparecían bien definidos en el citoplasma celular de la población, tanto de menor como de mayor tamaño (Figura 2a). Por el contrario, un mínimo porcentaje (0,98%) de la población celular presentó expresión para el marcador alfa-actina. Los filamentos de alfa-actina aparecían bien definidos en la población celular de mayor tamaño, en la que la actina se encontraba polimerizada en filamentos (Figura 2b). En el resto de la población, podía observarse en algunas células un marcaje punteado en el citoplasma que indicaba la existencia de actina sin polimerizar. Cuando el marcador CD31 fue analizado, un mínimo porcentaje de la población celular obtenida del pterygium (0,39%) mostraba expresión (Figura 2c).
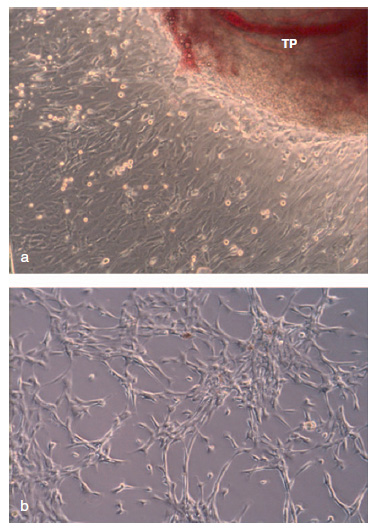
Figura 1. a) Imagen de microscopia óptica invertida de un explanto
de tejido obtenido del pterygium (TP), a partir del cual se obtienen
las poblaciones celulares en cultivo (10x). b) Aspecto del cultivo
celular tras el primer subcultivo (10x).
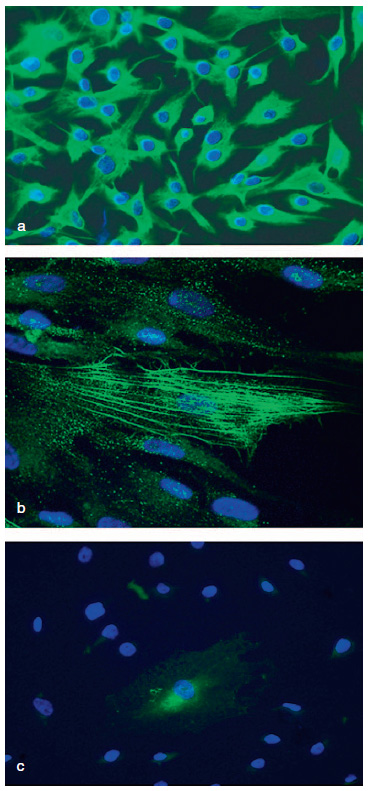
Figura 2. Imágenes de las poblaciones celulares procedentes
de pterygium sometidas a ensayos de inmunofluorescencia con
anticuerpos primarios: a) anti-vimentina (20x), b) anti-α-actina
de músculo liso (63x), y c) anti-CD31 (20x). Todos los núcleos
celulares se encuentran contrastados con DAPI.
En el análisis de la expresión de los marcadores de células madre embrionarias se comprobó que la mayoría de la población celular era positiva para ambos marcadores, OCT3/4 y CD9. El marcaje para OCT3/4 aparecía en todos los casos localizado en diferentes áreas específicas dentro del núcleo celular. Prácticamente la totalidad de la población celular (97,31%) mostró expresión positiva para OCT3/4 (Figura 3a). Por el contrario, el marcaje para CD9 se localizaba a nivel de la membrana plasmática, sobre toda su superficie. La totalidad de las células obtenidas del pterygium mostraron positividad para CD9 (Figura 3b).
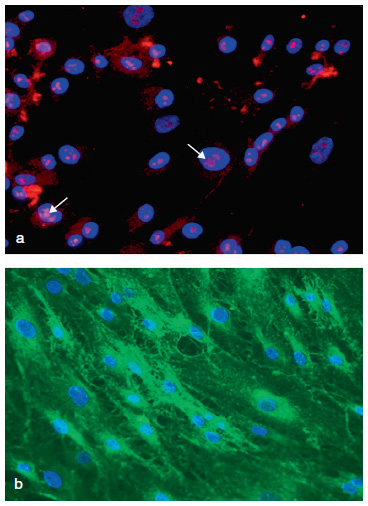
Figura 3. Imágenes de la identificación con técnicas de inmunofluorescencia
de los marcadores característicos de células madre embrionarias. a) Marcaje
para el factor de transcripción OCT3/4, limitado a zonas específicas dentro
del núcleo celular (flechas), (20x). b) Marcaje para CD9 (20x). Todos los núcleos
celulares se encuentran contrastados con DAPI.
Discusión
Clásicamente, el pterygium ha sido considerado como una enfermedad degenerativa crónica con degeneración elastoide causada por la irradiación ultravioleta11. Sin embargo, más recientemente se ha demostrado que el pterygium es en realidad una enfermedad proliferativa. Hallazgos histopatológicos revelan hiperplasia epitelial y un tejido fibrovascular exuberante en su estroma12. El objetivo de nuestro estudio ha sido caracterizar la población celular obtenida de este tejido y comprobar si expresan marcadores de células madre indiferenciadas.
No existe mucha bibliografía sobre el estudio de la población celular obtenida a partir de pterygium. En un estudio inmunohistoquímico13, se identificaron miofibroblastos contráctiles (α-actina positivos) en la población celular obtenida in vitro, así como en el tejido extirpado de pterygium primario y recurrente. Nuestros resultados revelan que en nuestra población, el porcentaje de células α-actina positivas es muy bajo (inferior 1%), mientras que se expresa mayoritariamente vimentina, La vimentina es un filamento intermedio del citoesqueleto, expresado por las células indiferenciadas en cultivo para adherirse y adaptarse al sustrato subyacente. La expresión de vimentina ha sido referida por varios autores13,14 en su población de miofibroblastos, afirmando que el pterygium se originaría por la invasión de células basales epiteliales limbales alteradas que expresan vimentina. Además, se ha observado la presencia de células epiteliales que expresan vimentina no sólo en la superficie del pterygium, sino también en la conjuntiva aparentemente normal adyacente al pterygium, sugiriendo que la resección quirúrgica incompleta de estas células sería la responsable de la alta tasa de recurrencia del pterygium15.
Dado que la expresión de vimentina se relaciona con células en un estadio de indiferenciación, nos propusimos investigar la expresión de marcadores de estadios más precoces en la diferenciación celular. Existe un gran número de factores de transcripción esenciales para el mantenimiento del estado de las células madre embrionarias, pero el OCT3/4 es el mejor caracterizado. El OCT3/4 es esencial para el mantenimiento de la pluripotencialidad de diferentes tipos celulares, está críticamente implicado en la autorrenovación de las células madre embrionarias indiferenciadas y debe ser expresado por encima de un nivel crítico para preservar el estado de célula madre16.
Este estudio utiliza marcadores de células madre indiferenciadas, como son el OCT3/4 y CD9, con el objetivo de caracterizar el grado de diferenciación de la población celular obtenida de pterygium. El marcador CD9 es una proteína transmembrana que se expresa en células madre embrionarias indiferenciadas y disminuye su expresión significativamente tras la diferenciación celular17. Nuestros hallazgos revelaron que los marcadores fueron expresados prácticamente por la totalidad de la población, sugiriendo que estas células estarían en un estadio de diferenciación más primitivo que el referido por otros autores. Otros estudios inmunohistoquímicos18-21, utilizando marcadores de células madre adultas, como C-Kit, CD34, STRO-1 y AC133, han revelado que existen células multipotentes en el pterygium que estarían involucradas en la patogénesis, progresión y recurrencia del mismo.
Así pues, la población celular expandida a partir de pterygium está compuesta por células de fenotipo mesenquimal que muestran expresión de marcadores embrionarios. Estos hallazgos nos sugieren que pequeños fragmentos de pterygium remanentes tras la extirpación quirúrgica podrían comportarse de un modo similar al cultivo, pudiendo por tanto, esta población celular ser la responsable de la alta tasa de recurrencia de este tejido tras la cirugía.
Conflicto de intereses
Los autores declaran no tener ningún conflicto de intereses.
Bibliografía
1. Jaros P.A., DeLuise V.P. Pinguecula and pterygia. Surv Ophthalmol. 1988; 33:41-9. [ Links ]
2. Hill J.C., Maske R. Pathogenesis of pterygium. Eye. 1989; 3:218-26. [ Links ]
3. Cameron M.E. Histology of pterygium: an electron microscopic study. Br J Opthalmol. 1983; 67:604-8. [ Links ]
4. Tsai Y.Y., Cheng Y.W., Lee H., Tsai F.J., Tseng S.H., Lin C.L., et al. Oxidative damage in pterygium. Mol Vis. 2005; 11:71-5. [ Links ]
5. Shimmura S., Ishioka M., Hanada K., Shimazaki J., Tsubota K. Telomerasa activity and p53 expression in pterygia. Invest Ophtahlmol Vis Sci. 2000; 41:1364-9. [ Links ]
6. Dushku N., John M.K., Shultz G.S., Reid T.W. Pterygia pathogenesis: corneal invasion by matrix metalloproteinase expressing altered limbal epithelial basal cells. Arch Ophthalmol. 2001; 119:695-706. [ Links ]
7. Van Setten G., Aspiotis M., Blalock T.D., Grotendorst G., Schultz G. Connective tissue growth factor in pterygium: simultaneous presence with vascular endothelial growth factor, possible contributing factor to conjunctival scarring. Graefes Arch Clin Exp Ophthalmol. 2003; 241:135-9. [ Links ]
8. Solomon A., Grueterich M., Li D.Q., Meller D., Lee S.B., Tseng S.C. Overexpression of insuline-like growth factor-binding protein-2 in pterygium body fibroblasts. Invest Ophthalmol Vis Sci. 2003; 44:573-80. [ Links ]
9. Marcovich A.L., Morad Y., Sandbank J., Huszar M., Rosner M., Pollack A., et al. Angiogenesis in pterygium: morphometric and immunohistochemical study. Curr Eye Res. 2002; 25:17-22. [ Links ]
10. Lee D.H., Cho H.J., Kim J.T., Choi J.S., Joo C.K. Expression of vascular endothelial growth factor and inducible nitric oxide synthase in pterygia. Cornea. 2001; 20:738-42. [ Links ]
11. Austin P., Jakobiec F.A., Iwamoto T. Elastodysplasia and elastodystrophy as the pathogenic bases of ocular pterygia and pinguecula. Ophthalmology. 1983; 90:96-109. [ Links ]
12. Tan D., Chee S.P., Dear K., Lim A.S. Effect of pterygium morphology on pterygium recurrence in a controlled trial comparing conjuntival autografting with bare sclera excision. Arch Ophthalmol. 1997; 115:1235-40. [ Links ]
13. Touhami A., Di Pascuale M.A., Kawatika T., Del Valle M., Rosa R.H., Dubovy S., et al. Characterisation of myofibroblasts in fibrovascular tissue of primary and recurrent pterygia. Br J Ophthalmol. 2005; 89:269-74. [ Links ]
14. Dushku N., Reid T.W. Immunohistochemical evidence that human pterygia originate from an invasión of vimentin-expressing altered limbal epithelial basal cells. Curr Eye Res. 1994; 13:473-81. [ Links ]
15. Chen Y.T., Tseng S.H., Tsai Y.Y., Huang F.C., Tseng S.Y. Distribution of vimentin-expressing cells in pterygium: an immunocytochemical study of impression cytology specimens. Cornea. 2009; 28:547-52. [ Links ]
16. Niwa H., Miyazaki J., Smith A.G. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat Genet. 2000; 24:372-6. [ Links ]
17. Aoyama K., Oritani K., Yokota T., Ishikawa J., Nishiura T., Miyake K., et al. Stromal cell CD9 regulates differentiation of hematopoietic stem/progenitor cells. Blood. 1999; 93:2586-94. [ Links ]
18. Ye J., Song Y.S., Kang S.H., Yao K., Kim J.C. Involvement of bone marrow-derived stem and progenitor cells in the pathogenesis of pterygium. Eye. 2004; 18:839-43. [ Links ]
19. Song Y.S., Ryu Y.H., Choi S.R., Kim J.C. The involvement of adult stem cells originated from bone marrow in the pathogenesis of pterygia. Yonsei Med J. 2005; 46:687-92. [ Links ]
20. Lee J.K., Song Y.S., Ha H.S., Park J.H., Kim M.K., Park A.J., et al. Endothelial progenitor cells in pterygium pathogenesis. Eye. 2007; 21:1186-93. [ Links ]
21. Lee J.K., Kim J.C. Progenitor cells in healing after pterygium excision. Yonsei Med J. 2007; 48:48-54. [ Links ]
 Dirección para correspondencia:
Dirección para correspondencia:
cinta.perezrico@gmail.com
(C. Pérez-Rico)
Recibido 24 Julio 2010
Aceptado 10 Noviembre 2010

