










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkInternational Microbiology
versão impressa ISSN 1139-6709
INT. MICROBIOL. vol.8 no.2 Jun. 2005
| RESEARCH REVIEW | |||
|
| |||
| Evolution and the nature of time
Summary. The concept of time is critical in evolutionary thought, but rarely has it been considered as an object of theoretical research by evolutionary biologists. Evolution is an organism's possibility of access to the future; in other words, evolutionary reward is paid out as increased time. Replicating entities are granted time, but for them, time only serves to allow replication and evolution, and to further expand the frontier of time. The present review discusses the possible influence of considering time not as a pure dimension (or an a priori intuitive condition of human experience) but as an object in itself. At least as a metaphor, time can be considered as a self-replicating entity rooted in physical (including biological) beings, with the result of producing dimensional time. Time self-replication forces beings to replicate, which, in turn, further sustains the replication of time. In that sense, time-replication may constitute the driving force, i.e., the basic engine, providing directional energy to the evolutionary process. The philosophical roots, caveats, and perspectives of this hypothesis are presented here. The metaphor of replicating-time plays with the possibility of viewing time not as a merely regulatory component of scientific inquiry but instead, as a real and creative constituent of nature and, for this reason, an object worthy of research in the natural sciences. [Int Microbiol 2005; 8(2):81-91] Key words: evolution · evolutionary time · time (as a replicating entity) · time (nature) | ||
|
| |||
Evolución y naturaleza del tiempo Resumen. En el pensamiento evolutivo, el concepto de tiempo es crítico, pero los biólogos evolutivos raramente lo consideran como un objeto de investigación teórica. La evolución significa una posibilidad de acceder al futuro de los organismos replicativos; es decir, la recompensa evolutiva se paga con la extensión del tiempo. La entidades replicativas obtienen tiempo, pero para ellas el tiempo sólo sirve para replicarse, para evolucionar y para expandir más allá la frontera del tiempo. Revisamos y tratamos aquí la posible influencia de considerar el tiempo, no como una mera dimensión (o una condición intuitiva a priori para la experiencia humana), sino como un objeto en sí mismo. Al menos como una metáfora, podemos considerar el tiempo como una entidad autorreplicativa enraízada en los seres físicos (entre ellos, los biológicos) que acaba produciendo tiempo dimensional. La autorreplicación del tiempo fuerza a los seres a replicarse para sostener la subsiguiente replicación del tiempo. En este sentido, el conjunto tiempo-replicación puede constituir la fuerza motriz, el motor básico que proporciona energía direccional al proceso evolutivo. Se presentan aquí las raíces filosóficas, los caveat y perspectivas de esta hipótesis. La metáfora del tiempo replicativo juega con la posibilidad de devolver el tiempo desde su función meramente reguladora de la investigación científica a una posición de constituyente de la naturaleza real y creativo y por ello objeto de investigación para las ciencias naturales. [Int Microbiol 2005; 8(2):81-91] Palabras clave: evolución · tiempo evolutivo · tiempo (como entidad replicativa) · tiempo (naturaleza) | Evolução e natureza do tempo Resumo. No pensamento evolutivo, o conceito de tempo é crítico, mas os biólogos evolutivos raramente o consideram como um objeto de pesquisa teórica. A evolução significa uma possibilidade de acessar ao futuro dos organismos replicativos; ou seja, a recompensa evolutiva é paga com a extensão do tempo. A entidades replicativas obtêm tempo, mas para elas o tempo só serve para se replicar, para evolucionar e para expandir além a fronteira do tempo. Revisamos e tratamos aqui a possível influência de considerar o tempo, não como uma mera dimensão (ou uma condição intuitiva a priori para a experiência humana), mas como um objeto em si mesmo. Pelo menos como uma metáfora, podemos considerar o tempo como uma entidade autorreplicativa enraizada nas naturezas físicas (entre eles, os biológicos) que acaba produzindo tempo dimensional. A autorreplicação do tempo força as naturezas a se replicar para sustentar a subseqüente replicação do tempo. Neste sentido, o conjunto tempo-replicação pode constituir a força motriz, o motor básico que proporciona energia direcional ao processo evolutivo. Se apresentam aqui as raízes filosóficas, as advertências e as perspectivas destas hipóteses. A metáfora do tempo replicativo joga com a possibilidade de devolver o tempo desde sua função meramente reguladora da pesquisa científica a uma posição de constituinte da natureza real e criativo e por isso objeto de pesquisa para as ciências naturais. [Int Microbiol 2005; 8(2):81-91] Palavras chave: evoluçao · tempo evolutivo · tempo como entidade replicativa · tempo (natureza) |
Die Zeit ist die Unruhe des Seins
Anton Neuhäuser, Zeit und Sein, 1957
Il est vrai qu'on n'est pas d'accord sur ce qui se gagne ni sur ce qui se perd entre le tour de la naissance et celui de la mort
Henri Bergson, L'évolution creatrice, 1907
Time and directional change in evolution
The fundamental question in evolution remains largely unexplored and, consequently, unanswered: What constitutes the driving force leading organisms to evolve in particular directions? Obviously, the question whether or not there is a direction (note that we are not mentioning progress) in biological evolution is a matter of controversy [37,46]. Elementary intuition suggests that there is a general trend of moving from simplicity to complexity. As Maynard-Smith pointed out, the first simple organisms had nowhere to go but up, to increasing complexity over time [39]. Indeed, the same intuition is at the bases of Spencer's law of evolution [49]. Complexity means intra-organismal complexity, as well as complexity in number, sort, and type of organisms, and complexity in the associations of organisms. Indeed, complexity means an association and implies a structured interaction [9,10].
At first sight, this tendency to increasing biological complexity could correspond to the analogous tendency to increasing complexity (from the random to the structured) that is also found in inanimate matter, probably including the entire expanding Universe. As Spencer suggested a long time ago [50], since living beings only exist (survive) if they constantly adapt to their environments, the driving force of the changing forms of life may be changes in their inanimate environments-non-living wheels transmitting movement to the living wheels in the universal clock. Changes in the complexity of the inanimate world produce changes in the structure of biological beings. At second sight, we realize that the correspondence between environmental and biological changes may not be linear. For instance, the speed of biological evolution seems to increase during those periods in which there are no (apparent) major changes in the evolution of inanimate matter. In that case, an effect based on the relative sizes of the clock wheels could be taking place; that is, high-speed evolution in the small biological wheel could be needed to adapt to low-speed, but significant evolution in the large inanimate wheel. In microbiology, there are some experimental examples in which very small differences created along gradients emerging in inanimate environments create significant changes in selection and presumably in evolution [7,8]. There is also evidence that evolutionary rate and genetic complexity may accelerate in particular environments [11,42]. At third sight, we could arrive at a trivial notion, i.e., biological evolution might accelerate independently from non-biologically driven changes in the inanimate world, because life is self-catalyzing: the greater the variety of life forms, the greater the variety required to adapt to this variety. This hypothesis has been studied by Waddington [53].
Such initial reflections do not help to provide any light on the first basic question: what constitutes the driving force, the basic engine providing directional energy to the evolutionary process? The notion of evolution was born in relation to the struggle for life. The action of the living being (any unity of selection) is confronted with the reaction of the environment, and evolution enables the organism to survive. Certainly, the basic question can be also stated in this way: what explains the (apparent) intrinsic tendency of living beings to evolve in order to survive? How does a given direction of the time-arrow influence currently living organisms? How may the presumptive benefits of a future life decide (or model) the behavior of present organisms? In other words, how could the future act as an attractor driving the dynamics of the evolutionary system? In the old terms that Emmanuel Kant used to define biological matter [32], is there a purposive mechanism of evolution? We can simply answer that the last three questions are false questions, concluding that, in biology, any notion of purpose is just an anthropic illusion. Evolution could eventually be conceived as a purposeless effect, a passive, and not an active attribute of biological beings. In fact, there are no winners in the struggle for life, but just blind (malgré soi) survivors of environmental changes. But, in any case, the main, almost metaphysical question still remains: why do biological beings tend to replicate? Why feed a system devoted to survival and preservation of a particular biological identity? What fuels evolutionary biology?
Why replicate?
We should consider that the maintenance of biological identity cannot be identified as the evolutionary goal, because such self-fidelity may not be preserved over long-term evolution. Most of us agree that, considering a given (short) period of time, species of organisms may be well described as discrete phenomena of nature, tending to protect the integrity of a particular gene pool [40]. However, if very long periods of time are considered, the once harmonious gene combination may decrease in competitive effectiveness and, therefore, may be modified by evolution [10]. To solve this "species problem," it has been postulated that the identity (specificity) of organisms should be conceived along the time-arrow. In this way, species are spatio-temporal units [22,31]; that is, "evolutionary species" [48] having "historical essences" (Sykora 1995, personal communication). We can imagine these time-dimensioned species, or chronospecies, as timeworms, essentially uniform cylinders aligned along the temporal dimension. They have their counterparts in the more conventional space-dimensioned species, which could be called topospecies. The possibility of reconciling such (respectively) essentialist and nominalist approaches will probably arrive from a future consideration of the object "species" in a four-dimensional space-time frame! A systematic analysis of all these possibilities was presented by Stephen J. Gould, in the light of the hypothesis of hierarchical evolution, discussed in his last book, The Structure of Evolutionary Theory [27].
The essential component of the evolutionary process appears to be maintenance of the ability to replicate [41]. To exist is to survive, as punctual existence is nothing. To survive is to replicate, and hence to replicate is to exist. Both the term and the concept of survival (super-vivere) mean ‘going beyond the current life span, so that a certain trend to gain the future (a desire, again in Kant's terminology) seems to be deeply ingrained in biological matter. As Jorge R. de Santayana said, repetition is the only form of permanence that nature can achieve. In the words of Bergson, the persistence of life consists in a series of palpitations [12]. But, again, what constitutes the driving force for such an interest in the future? The historical fate of an organism is actually nothing, and it is impossible to believe that this absence can attract anything real. Therefore, if external influences are discarded, then the driving force should be internal to the organism, an apparently intrinsic property linked to biological matter.
Returning to the classic metaphor of the biological clock, we could imagine that there are minuscule energetic wheels in any living organism, transmitting after each round its obsessive, fractal message to the whole organism: you replicate as I am replicating. An idea of that kind probably led Richard Dawkins's to propose his famous "selfish genes" theory [16]. Replicators are able to influence the production of survival machines for themselves. Is gene replication the driving force of evolution? Even if it were so, the basic question remains again unanswered, perpetuates throughout the different biological levels, and is increasingly difficult to answer: what forces a gene to replicate, to survive? Are genes, and in general replicators, working for some other element, the very last selfish element?
Where is the profit of the evolutionary process? Evolution is based on profit, but for whom is this profit intended? Species are either modified over time or eliminated; individual organisms disappear; gene pools are shuffled by horizontal gene transfer. Is the profit for Life an abstraction? If so, under which terms would an evolutionary benefit be obtained? At this point, the answer is relatively simple: replication and survival of all biological organisms only serve to gain space and time. Simplifying Wilson's distinction [55], gain in space is dominance, whereas gain in time is success. Indeed, gain in space facilitates the settlement of more replicating entities and assures a supply of energy for replication. As the only sense of replication is to gain a future, finally, all evolutionary rewards are paid with more time. By replicating, organisms obtain time. For them, however, time only serves to allow replication, to evolve, and to further expand the frontier of time. Modern artificial-life evolutionary studies use digital organisms [1,56]. These are self-replicating computer programs that compete for central processing unit (CPU) time, which is the fuel needed for their replication [33]. Evolutionary reward is paid in expanding time. In a sense, time is the only selector; only if a given instant of time is surpassed, is another bit of time provided to the organism. In other words, time is both the selector and what is selected; organisms are, in this sense, co-selected with time, and pushed in such a way into the evolutionary road.
Is time something other than a dimension?
In this theoretical context, we could try to imagine time as nothing more than a cold pure dimension, as a Kantian a priori intuitive condition of experience, or as a framework for the subject's power of representation. Let us consider, at least as an exploratory metaphor in evolutionary theoretical research, time as something more than an object only in appearance, but as a thing, a true object. The nature of this time-object may contribute to the emergence of new events. It is difficult to conceive of an evolving nature as based only in physics or chemistry, whose laws tend to assure symmetry and equilibrium. But what if we consider time as an objective component of changing material objects, forcing them to change? It is the asymmetrical nature of time [28] that infects material objects and creates an evolutionary asymmetrical trend. To explain its asymmetrical nature, time may be well conceived as the ultimate self-replicating entity-in a broad sense, as a replicator; that is, "anything in the universe of which copies are made" [17].
Now we should approach the possibility that time needs matter to replicate within. Time cannot exist without matter. Its growth depends on the evolutionary success of time-expanding replicating entities, in our case, biological individuals. This means that time may be imagined as a parasite of biological matter. A parasite that, like other parasites, expands at the expenses of its host's matter and energy In the host, the end of time is death, which occurs when the parasite has totally exhausted the host's biological energy. By contrast, the successful survival and replication of the host assures time's replication, which forces the host to change and adapt, thereby fueling the evolutionary process. Selfish time transmits an obsessive signal: you replicate as I am replicating. Survive and evolve to replicate. Everything replicates for the profit of time, but only time replicates for its own profit. The nature of time is only to replicate, but doing so may prime the basic engine of evolution. Maybe the best expression of the signal is; reproduce as I am replicating. The wise differentiation between replicators and reproducers, coined by Szathmary and Maynard-Smith [51], may also be applied here, at least as a general concept. In our theoretical context, if time is the ultimate replication entity, able to produce an unlimited heredity or repeated units, then organisms are simply reproducers, able to expand time after time. Evolution assures the permanence of organisms, the real time-machines. They serve not to travel in time, but to produce time, to get more and more organisms, and thus more time, and thus more evolutionary time.
The hypotheses of a directional arrow resulting from natural time replication in biological matter could help to explain the directional behavior of the evolutionary process. In fact, evolution means directional orientation of biological changes [25]; in our hypothesis, the essence of the evolutionary process is to produce time with time. The emphasis on survival (necessarily of the fittest, again a conventional tautology) in evolutionary thinking expresses only an operative aspect of the process. The survivor is merely the organism that produces time; indeed it is a survivor because it produces time. Therefore, it is the most evolved, which means the most advanced of its lineage in time-production. Let us imagine that the time required to build and to maintain a non-replicative organism vanishes after its individual life span. The energy invested in expanding time is lost through the process. By contrast, evolutionarily successful replicating organisms contribute in an important way to spreading time.
The metaphor that is examined in this review is, like any other metaphor, necessarily speculative, or at its best, based on some philosophical arguments and not on particular facts. But there is no such a thing as philosophy-free science [18], and purely mental exercises on basic problems are required from time to time. As was stated a few years ago, the fact of the matter is that we simply cannot escape metaphorical thinking in science [19].
The nature of the time-dimension and the nature of time as a replicating entity
The philosophical question underlying considering time as a replicating entity is undoubtedly the definition of time. There are at least three possibilities of how to conceive of time; (1) as an immobile "cosmic wastebasket" sitting there waiting for events to be deposited into it; (2) as a dimension that serves to manifest relations between events, including changes (durations); and (3) as an object (thing) in itself. Since everything we know, including ourselves and also our thoughts, is time-contaminated, we have the innate (anthropic) tendency to consider time as an absolute dimension. Because of its apparent universality, time serves in practice to establish correspondences among those types of events that influence all aspects of our experience. Indeed, time, as space, has been considered as an intuition with the dynamic function of positing a manifold of intuitions as appearances. The experience and the consequent description of life, and certainly of evolution, is entirely dependent on time as a formal element of intuition. John A. Wheeler proposed the concept of "observer participancy description" of the universe, in which the creator of time is the human observer [54]. From Kant's point of view, time by itself is not an apprehensible object, as it is not an object of perception whose systematic connection can be termed experience. This concept became an obsession for Kant during his last days, and he re-iterated it many times in his dramatic last notes on the natural sciences [32]. As in the case of the definition of the color green, anybody intuitively knows what green is, but nobody can relate it to someone else's experience about what is green (at least, the green feeling), without resorting to the tautology "green is the color of green things". Beginning with Aristotle, time has been conventionally considered as the distance between two events: time is the measure of change with respect to before and after. As in the case of green, this definition is of extreme logical weakness, because its main conceptual components-change, before, and after-tautologically imply the defined time. The same is true if we use the word "event" in the definition. Can time be defined in the absence of time-based concepts? Mono-dimensional space may be defined as the distance between two points. Here, distance implies space again. Because of these difficulties, time and space were defined by Kant as a priori conditions of experience. The application of anthropic principle justifies Kant's conclusion: since humans are located in this particular time-frame, everything will be understood in the same time dimension. If we consider time or space as mere dimensions, it is certainly tremendously difficult to reach an accurate definition.
Let us now examine again the possibility of defining time as a naturally replicating object by proposing that time is a self-replicative natural entity rooted in matter; as a consequence of time's replication, matter changes. Self-replication should be interpreted here in the sense of autopoiesis [52]. Initially, this seems again to be a tautology. Nevertheless, this definition includes: (i) the concept that time is a natural element (entity); (ii) that itself-replicates; (iii) that it is matter-dependent; and (iv) that the change in the dimensional time of matter depends on it. In order to eliminate the tautology, we need to differentiate between naturally replicating time (time-element; the replicating entity) and dimensional time, in which the time-element is converted (to a certain extent, expressed).
Obviously, the major scolio in our time-as-a-replicating-entity metaphor is the nature of the thing that the time-element (as an object) actually is in the objective world. Without continuous conversion into dimensional time, difficult for us to imagine how the time-element functions as a replicating entity (implying ancestors and successors). Indeed, the procession from ancestors to successors implies time; the time-ancestor in replication needs time to give rise to its time-successor. We could imagine that the time-replicating entity is constantly transformed into a time-dimension, enabling replication itself. With this conversion, it is possible to imagine time as consisting of elementary "chronons", comprising ancestors and successors, and that each element-successor is able to eliminate its corresponding element-ancestor. This notion is reminiscent of Greek myths, in which intrinsic forces mediate the flow of time. This flow is made up of successive time generations. Succession by opposition is only possible if we can imagine time as having a discontinuous (generational) nature. One of the Greek expressions for time, aiwn, was probably coined from the root yu or ayu, expressing "source of life", and later "vital force" (the word youth, from the Latin juventus, is a derivative). Classic myths about time, in which Uranus, Chronos (Saturn), and Zeus, are the the major characters, have in common an internal father-son or son-father obsessive annihilation. This reflects a very accurate intuitive representation of the essentially sequential nature of the flow of time. Each father-instant replicates in a new son-instant that kills its father. In neoplatonic philosophy, and particularly among the orphic fragments, an obscure time-god entity (Chronos) is primarily responsible for the unfolding of the universe (the Hegelian Entwicklung) from the primordial egg; that is, time as a primary engine. Perhaps this was an ancient way of expressing the following concept. Because of its nature as a primary replicating entity, time generates order (including the time dimension) from a previously random, chaotic world. It may be that order itself is (or was) time-dependent. In other words, the selective advantage of order over chaos in spatially extended systems may consist of its ability to replicate because of time.
To facilitate an intuitive appreciation of the possible nature of biologically produced time-element, we could propose a comparison with another well known biologically produced derivative, such as heat (calories). As in the case of time, heat is produced by replicating organisms and is dissipated by death. For obvious reasons, heat (the fire of Greeks) was classically considered to be the soul of animal matter. Forcing the analogy with time, we could imagine that heat is a replicating entity, a parasite of matter (as there is no heat without matter), and also a parasite of living beings, forcing them to replicate and to evolve to increase the quantity of biologically produced heat. Everything about our heat hypothesis looks similar to the time-replicating hypotheses, except for a major point; Heat is not directional, but dispersive, and there is not a heat-arrow exerting directional pressure.
Time in evolution: time for event formation and evolution
In considering time as a replicating entity, matter is contaminated with a time-parasite, and the time dimension begins to flow. Obviously, no time exists without matter, as no strict parasite exists without a host. A corollarium of this concept is that matter should precede time, and we must admit the possibility of a time a-dimensional material world. Thus, the time-replicating entity would have emerged as a parasite only (at random) in certain parts (singularities) of a previous simultaneous universe. If that were the case, then a-temporal matter could exist even today. By applying classical means of mental representation, time could be also conceived as the anima, the changing-moving force, and as the soul of matter. In this perspective, the expression "inanimate matter" is much more restrictive, describing only the presence of matter before time, in a purely spatial universe. The linkage between time and matter was also a classic theological question: "What did God do before creating the universe?" But here, in the internal logic of theology, "before" has no sense. Saint Augustine considered time as non-existing before the beginning of matter, and a property of the created universe [3; implicitly in Confessions, IV: 11]. In a certain way, that reflects the idea of the matter-dependent nature of time, if not of the material nature of time itself.
Applying our metaphor to biological matter, we have suggested that time produces time through the intermediate of organismal replication. In this case, the time-replicating element might be called biologically generated time, and the result is visible as dimensional time, which may be conceived as the phenotype of replicating time. In our model, the time-replicating entity generates dimensional time for event formation. The signal for the birth of a new organism may have originated because the time-replicating entity was replicating; but once that occurs, the organism has time. In other words, it receives the time-heritage (or time-parasite) of its ancestor.
The acquired time is converted into dimensional time, and enables the organism to cultivate those events that assure multiplication and hence the transmission of its time-replicating entities to its successors. If replication does not take place, time is lost. Once the host organism is exhausted by time, its time is dissipated. Species with very low numbers of rarely reproducing individuals are at risk of extinction because time is at risk of being dissipated in these species, impeding the event processes of adaptation. By contrast, species with high densities of individuals and high replication rates (such as bacterial species) produce huge densities of time for event formation and evolution. This point is discussed below.
Event formation in the biological and physical world
A correspondence can be found between the driving role of time in the production of events (evolution) in the biological and the physical (cosmological) world. This does not imply that the presumed object "time" is necessarily the same in both contexts. One of the advantages of the liberation of the concept of time from its absolute character of ens rationis is the possibility of considering the heterogeneity of time. Different types of time in different places, perhaps with different replication rates according to the different material hosts, suggest different tempus and modes in branching evolution. The discussion on the influence of time in the Entwicklung of the physical world is beyond the scope of this work. But the key facts resulting from the theory of relativity, i.e., the existence of a limit in time-speed and the modifications of time-flow by physical force fields (for instance, gravitational), as well as the problem of simultaneous measurements, leading to the principle of indeterminacy, tend to support the non-purely dimensional abstract nature of time. The theory of quantum gravity indeed suggests that time is constructed from discrete elements [14]. In any case, it is curious that the concepts of quantum physics have contributed very little to biology.
As Stephen Hawking said, time not only affects but is also affected by everything that happens in the universe [30]. The controversial part of this sentence is how time affects, or is affected. Time as a dimension cannot affect, or be affected by anything, just as a y-axis cannot affect the coordinates of any point, or as the coordinates cannot affect the axis. The y-axis by itself is absolutely nothing but a virtual line to place particular values on. The values define the coordinate, not the axis. Particular events (placed in a time t-axis) modify reality, but not the dimension of time itself. The axis of time cannot modify events. If we seriously say that time affects, or is affected by, objects in the universe, we are implicitly saying that time is an object, and not a dimension. If that is true, then how can time be differentiated from other objects in nature?
According to the view of Einstein, time (space-time) has an built-in tendency to expand. How can an axis, the t dimensional axis, expand, if time is only an entity of pure reason? Only things can expand; the expansion of mentally derived entities can be better defined as imagination. Similarly, how can space-time attract the matter of the universe without being itself an object? In more biological terms, how can time affect a physiological process such as aging? Coming back to the concepts of Kant, maybe time does not affect physiological processes at all, and the expression "time affects" is only a subjective, mental representation of a directional series of phenomena occurring in biological organization. This view maintains the mystery of what on Earth may produce the directional, sequential series of changes that characterize the evolutionary process. But we can imagine that there is a time that objectively affects biological processes, acting as a real force and mediated by the activity of real, objective elements. The concept of space-time as a mere receptacle in which physical phenomena happen may change to one depicting space-time as the origin and driving force of the evolution of the physical world itself. This is the metaphor that we are exploring here.
Replicating time: the time arrow
Conventional dimensional time is considered to be uniform and constant over time [43], again a self-referenced definition, as the intellect is "accustomed to think the moving by means of the unmovable" [12]. How is it that time might change without other standards of time that can be used to measure it? That is another mystery of the sort with which mathematics is strangely replete (13). The hypothesis of time as a replicating entity may allow an escape from such logic, by emphasizing the distinction between replicating time and time as a dimension. The distinction between these two kinds of times was proposed by Newton in his Principia, which described a real absolute time, that "flows by its own nature"; and a "relative" time (or duration), an ens rationis, that serves only to measure and that "is frequently used instead of the real time". The time arrow (and its projection, eternity) is a anthropic image of perceived relative or dimensional time, but it cannot be applied to fragmented, replicating natural time, which depends on matter. If dimensional time results from natural, replicating time, and if natural time is matter-dependent (because every parasite needs a host), there is the possibility of a heterogeneity in the time arrow. This possibility has been already mentioned by others [24,25]. In a sense, time arrows point to different directions, changing in trajectory, but in all cases they are directionally and irreversibly oriented . In other words, they resemble natural objects in evolution.
There is an intuitive attraction of biologists, influenced by the observation of the apparent cycles of reproduction of living organisms, to the cycling of time [35]. The possibility of a time cycle (moving from the present to the past, in the Newtonian view of time) will be difficult or impossible to reconcile with any autopoietic view of time producing time. A replicating entity produces a progeny, which tends to separate from its ancestors. This separation is essentially due to dimensional time being generated by replicating time; to a certain extent, the interposition of time is causes increased distance as well as divergence between members of the progeny. In general, we can imagine that dimensional time spread produces a (physical?) separation between time-replicating elements and their offspring.
It is intuitively clear that any arrow reflects a non-equilibrium system, perhaps pointing to a final equilibrium. The thermodynamic arrow of time is considered to be the direction of time in which disorder or entropy increases [15, 30, 36], which relates to the irreversibility of time. In discussing dissipative structures [44], Ilya Prigogine quoted Erwin Schrödinger: "There should be something in the mechanism of life that prevents life degradation, there should be a type of irreversible phenomenon" [45,47]. We could indeed consider a time-driven increase in biological expansion and complexity. Increasing complexity assures the diversification of beings, and consequently survival in the form of the accretion of replicating entities and time expansion. The cosmological arrow of time refers to the direction of time in which the universe expands. Again, in the time-replicating view, the expansion of the universe might be considered as triggered by this replicating time (but we leave the discussion of this point to physics). The only possible arrow resulting from the dynamics of time replication is the direction (probably, directions, or space) that the time progeny generates during its expansion. Like most replicating entities, at least during certain phases of growth, time might even expand exponentially. An example of this kind of growth is the documented accelerated expansion of the universe. But what is the limit then of the expansion of time? If the time-parasite is dependent upon its matter-host, the limit should be the extreme dilution of matter. The end of time will arrive when the concentration of matter at any location of the universe reaches zero; that is, the limit is the nothingness. Paradoxically, as in the case of parasite-prey interactions, time will collapse when after having reached its maximum dimension-unless, analogous to other parasitic replicating entities, a parasite (time)-prey (matter) equilibrium is reached. This implies that time only expands when a critical concentration of matter is available. We are far from converting our metaphor into a testable hypothesis, but these predictions may serve as the basis of a future model and experimental work.
Time and biological replicating units: interactive models
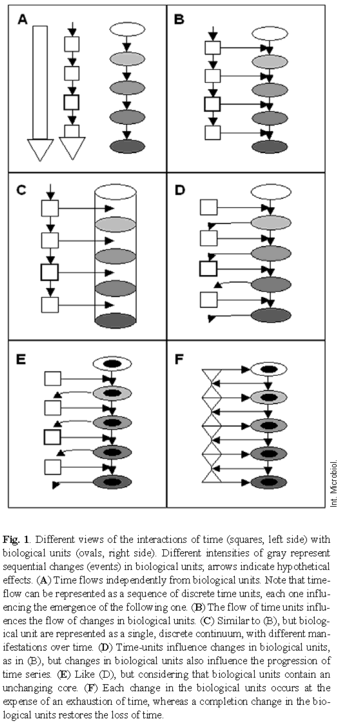
Figure 1 summarizes some of the models of possible interactions between time and biological replicating units. The first model (Fig. 1A) considers that replication, and hence evolutionary changes, in biological organisms are fully independent from the flow of time-that here, only dimensional time plays a role. Even if time could be considered as a chain of time-replicating units, they would be totally unrelated to biological activity The second model suggests that the flow of time-replicating units influences the flow of biological change, so that different time-inputs trigger biological flow-no matter that the organism is formed by discrete units limited in time (Fig. 1B) or as a trans-individual time-worm organism (Fig. 1C). The third model provides for the possibility that the accretion of time depends on biological replication or change: time-replicating units trigger biological replication, but this change also produces time, which is used to trigger the next biological change (Fig. 1D). Eventually, a part of the biologically replicating entity remains unchanged (Fig. 1E), in accordance with the theory of endurantism (Fig. 1F). In this third model, it could be considered that the process of transferring its replication energy to a biological unit exhausts each time-unit, but this energy is sequentially recovered as a result of successful biological replication and change (Fig. 1E).
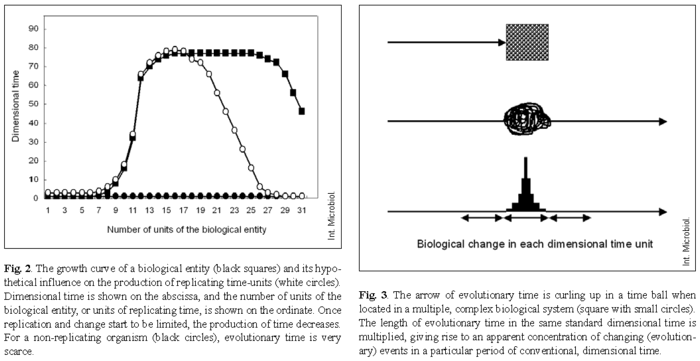
A necessary corollarium of this model is that there should be a local time, linked and interdependent upon local matter. Note that if the amount of time units produced by biological replication decreases, replication itself will decrease, so that, eventually, the local production of time and biological change will stop, resulting ultimately in death (Fig. 2).
The model of mutual interaction (interdependence) between the replication of time and that of biological matter can be expanded by considering that the biological units may interact with other biological units, whose replication will contribute also to the production of successive time-units. In this sense, any increase in the local density, variability, or connectivity of biological units is expected to result in a local increase of time. One may wonder whether the 3.85-billion-year period of conventional dimensional time, since the presumed origin of life on Earth, has been functionally equivalent for every type of living organisms. For instance, we could imagine that the absolute amount of cells, cell replication, cell differentiation, and cell interactions throughout the multiple environments colonized by the microbial world have multiplied the production of microbial time, so that, in fact, the evolutionary time for microbes has been by far much greater than that for other organisms. The events characterizing rapid evolution in certain periods (punctuated evolution) may not be more rapid in the absolute sense: rather, they accumulate when a high density and complexity of biological units favors the local production of evolutionary time, giving rise to increasing amounts of change and complexity. This concept is illustrated in Fig. 3.
Roots, caveats, and perspectives of the time-replication metaphor
The concept of time as an objective replicating entity is an extremely intuitive concept that was certainly present-but rarely recognized as such-in the minds of several philosophers, including Heraclitus and Zeno. According to the creationist perspective, time is a kind of "creature" (as proposed, for instance, by Maimonides), which implies a certain community with other more conventionally created beings. As E.H. Gombrich noted, the capacity of the human mind to defeat the flux of time and to perceive events that, strictly speaking, no longer exist provoked St Augustine to some of his profoundest meditations on the nature of time [23]. He realized than even in calling one syllable long and other short we are comparing sensations that had vanished (St Augustine, Confessions). The continuous presence of a given sensation could thus be interpreted as the persistence of some kind of physical "time-element". Henri Bergson [12] applied the same type of reasoning to evolution: "Evolution implies a real persistence of the past in the present, a duration which is, as it were, a hyphen, a connecting link". Leibniz, particularly after the Pacidius Philalethi (1676), considered all continuum, including time and space continua, as formed by discrete, substantial elements able to replicate harmoniously ad infinitum [20,29]. These monads certainly constitute one of the essential roots of our metaphor. Indeed, it was during the first quarter of the twentieth century that philosophers were closest to the intuition of time as a replicating entity. We should admit that our choice of the qualification "metaphor" to study the concept of time replication is based on what Bergson himself stated in 1911. "It is of the very essence of mechanism, in fact, to consider as metaphorical every expression which attributes to time an effective action and a reality of its own" [12]. Only occasionally does Bergson relate the "impetus that pushes life along the road of time" with time itself. But, at the end of his book Creative Evolution, the point is well-presented: "If time is not a kind of force, why does the universe unfold its successive states with a velocity which, in regard to my consciousness, is a veritable absolute? Why with this particular velocity rather than any other? Why not with an infinite velocity?". And then: "The more I consider this point, the more it seems to me that, [...] time taken up by this succession is something other than a number". And, a few pages later, he insists on the creative quality of time: "Time is invention or is nothing at all", and the "substitution of time-length for time-invention". These ideas were further developed in 1920, by Samuel Alexander, who probably offered a more relevant notion for examining the role of time as an engine in the evolutionary process. He considered time as composed of a continuum of substantial instants (events?), able to engender the processes modifying real objects [2]. In 1923, C. Lloyd Morgan developed the theory of emergent evolution, in which movement is an intrinsically emergent quality of time (or time-space) from which an impulse (nisus) is exerted on reality, forcing all beings to produce increasing complex (perfect) entities [34]. These ideas have been hidden from most evolutionary biologists. Interestingly, only someone from the fields of mechanics and thermodynamics, Prigogine, implicitly proposed time as a replicating entity of biological creatures. In his view, there is an "internal time" in the case of a chemical reaction, that dies when this reaction arrives at equilibrium. In the case of life, "an internal time is born, which is maintained along thousands of millions of years of life, being transmitted from one to other generation, from one to another species, and not only is it transmitted, but it increases its complexity" [45]. In even more recent times, Cahill and Klinger, working in process physics, have re-invented the concept of "pseudo-objects", which, like Leibniz's monads, have no intrinsic existence and are defined only by how strongly they connect with each other, serving as scaffolding to organize chaos into order. Steve Rosen has suggested that time might be self-generating in a life-similar process by self-referential transformations [Time Groups (2000) http://www.egroups.com/group/time]. Probably, in one way or another, from different angles and origins, the concept of time replication has begun to make its way into modern theoretical studies.
An obvious caveat for accepting the present metaphor about the role of time replication in driving the evolutionary process, is that our driver is too general to be seriously considered. By applying a Bergsonian view and acknowledging the complexity of the concept, time cannot be considered as an appropriate object for scientific study. Moreover, the Lamarckian notion of general "invisible fluids" (such as the river of time) acting on evolution nowadays seems unfashionable [37]. Classic ontological studies currently suffer from similar criticism; as studying the nature of being (ens) is too general to serve to explain anything. But, as ontology has served to fuel philosophical research, the hypothesis of time as a replicating entity may, at least, lead to a reconsideration of the role of time as an essential implicit component of evolutionary thought. Despite the view of some structural biologists [21], we should not seek to free ourselves from hypotheses-driven approaches, even if they are entirely theoretical. If we are interested in identifying the primary elements fueling Darwins's pump [18], it is not only the first "warm little pond" that should be investigated. Maybe the bases needed for understanding the origin of any evolutionary process are more deeply rooted than previously thought,. It is therefore unfortunate that the scientific community involved in evolutionary research has renounced theoretical investigations of time, one of the key words in evolution, but so rarely mentioned in the index pages of texts on evolution. In the years to come, increasing collaborations between physics and biology will make explorations of the natural world more efficient. What the Greeks called "metaphor" really means "transfer" or "carry-over". Like Prigogine [45], we believe that time is an object of scientific interest. Our replicating-time metaphor plays with the possibility of returning time from its merely regulatory function in scientific inquiry to a position as a real and creative constituent of Nature and, for this reason, an object of research for natural sciences.
Coda: time, evolution, and teleology
One of the major objections of Darwin to Lamarck's theories was their implicit notion that organisms exhibit an innate drive towards complexity, a teleological sense. As was recently discussed by Francisco J. Ayala [4], teleology describes actions, objects, or processes that exhibit an orientation towards a certain goal or end-state. In this sense, natural selection can be said to be a teleological process that results in increased reproductive efficiency, so that reproductive fitness is the end result or goal of natural selection. Our objection is that the production of time, rather than reproductive fitness may be the final end-state. Indeed the metaphor of time as a replicating entity may serve to better reconcile those views advocating that, ultimately, evolution has a meaning with those proposing that evolution is simply a meandering process going nowhere in particular. In our metaphor, the time-line does not precede the evolutionary process, rather time emerges simultaneously with the evolutionary progress line. In other words, the directional orientation of the process is linked to the last element in the progression. There is indeed a goal, to permit the replication of time, but it is not "external" to the evolutionary trend. And, simultaneously, there is no other direction than that of time replication, so that organismal evolution may appear to be going elsewhere, or even nowhere, but it is certainly not a random walk; it goes in the direction of time replication. In other words, the replicating-time metaphor is consistent with both internalist and externalist theories of directional evolutionary change [24,26].
Theoretical biology has always been envious of theoretical physics [6], in which apparently well-grounded, immovable basic principles sustain a geometrically structured universe. Nevertheless, in recent years, physicists have become suspicious of a fundamentally random universe, which has motivated exchanging geometric-static models of reality for biological ones. In this context, some research groups involved in process physics have started to model time not as geometry, but rather as a process, including that of self-replication. In their view, time and all of the objects around us, including those on which evolution is acting, are nothing more than the froth on a deep sea of randomness [14]. Time and objects in evolution may be linked as part of this advancing front. In his book Protobiology, Koichiro Matsuno [38] laid out the basis for a "physics of becoming" that s with the classical "physics of being". Conceptual biology has penetrated theoretical physics. Our metaphor, suggesting that time, which could well be acting as a replicating entity, expanding itself, and, as a result, fueling the evolving universe, may be considered as an example of this trend in the field of theoretical biology. As Prigogine said, to read the history of the Universe as the history of "autonomous time" is one of the most interesting temptations of contemporary science [45]. The role of science is to propose and discover links between natural objects, trying to overcome as much as possible, as Gould suggested [27], the parochial reasons of our personal inexperience. For that reason, and at least as a hypothesis, we should not exclude time, as a definitely creative force, from our considerations about the structural and evolutionary frame of Nature.
Acknowledgements. I acknowledge Stephen Paul King for his initial encouragement of an early draft of this manuscript. I also thank Marc Lipsitch (School of Public Health, Harvard University, Boston, MA), Richard Lenski (Michigan State University, East Lansing, MI), Bruce Levin (Emory University, Atlanta, GA), José Luis Martínez (National Center for Biotechnology, Madrid, Spain), and Kim Sterelny (Australian National University, Canberra, Australia) for useful comments and criticisms about the concepts of time in evolution that are discussed in this paper. Finally, this work would like to honor the memory of Stephen Jay Gould, who was, and remains, a source of continuous inspiration for scientists in a wide range of disciplines.
References
1. Adami C (1998) Introduction to artificial life. Springer, New York [ Links ]
2. Alexander S (1920) Space, time, and deity. The Gilford Lecture at Glasgow, 1916-1918. MacMillan, London [ Links ]
3. Augustine St. The literal meaning of genesis (JH Taylor, trans.) (1982) Newman, New York [ Links ]
4. Ayala FJ (1974) Introduction. In: Studies in the philosophy of biology. FJ Ayala and T Dobzhansky (eds) pp vii-xvi, Macmillan, London [ Links ]
5. Ayala FJ (1998) Teleological explanations in evolutionary biology. In: C Allen, M Bekoff, G Lauder (eds) Nature purposes: analyses of function and design in biology, pp 29-49, Massachusetts Institute of Technology, Cambridge, MA [ Links ]
6. Baquero F (1977) El método teórico en biología. Revista de Occidente 19:69-75 [ Links ]
7. Baquero F, Negri MC (1997) Selective compartments for resistant microorganisms in antibiotic gradients. BioEssays 19:731-736 [ Links ]
8. Baquero F, Blazquez J (1997) Evolution of antibiotic resistance. Trends Ecol Evol 12:482-487 [ Links ]
9. Baquero F, Coque TM, Cantón R (2003) Antibiotics, complexity and evolution. ASM News 69:547-552 [ Links ]
10. Baquero F (2004) From pieces to patterns: evolutionary engineering in bacterial pathogens. Nature Rev Microbiol 2:510-518 [ Links ]
11. Bastarrechea F (1997) On the origin of plasmid-borne, extended-spectrum, antibiotic resistance mutations in bacteria. J Theor Biol 190:379-387 [ Links ]
12. Bergson H (1907) L'évolution creatrice. (Electronic version available at the Paul-Emile-Boulet Library of the University of Quebec-Chicoutimi. [ Links ])
13. Berlinski D (1997) A tour of the calculus. Mandarin Paperbacks, London [ Links ]
14. Chalmers M (2003). Welcome to quantum gravity. Physics World, 11:27-50 [ Links ]
15. Davies P (2004) The quest for the source of the time arrow. Nature 428:257-258 [ Links ]
16. Dawkins R (1976) The selfish gene. Oxford University Press, Oxford [ Links ]
17. Dawkins R (1982) Replicators and vehicles. In: Current Problems in Sociobiology King's College Sociobiology Group, eds. pp 45-64. Cambridge University Press, Cambridge [ Links ]
18. Dennett DC (1995) Darwin's dangerous idea. Touchstone, New York [ Links ]
19. Draaisma D (2001) The tracks of thought. Nature 414:153 [ Links ]
20. Duchesnau F (1993) Leibniz et la méthode de la science. Presses Universitaires de France, Paris [ Links ]
21. Editorial (2000) Can biological phenomena be understood by humans? Nature 403:345 [ Links ]
22. Ghiselin MT (1974) A radical solution to the species problem. System Zool 23:536-544 [ Links ]
23. Gombrich EH (1979) The sense of order. Phaidon Press, London [ Links ]
24. Gould SJ (1977) Eternal metaphors of paleontology. In: Patterns of evolution. A Hallman (ed) pp. 1-26. Elsevier, Amsterdam [ Links ]
25. Gould SJ (1987) Time's arrow, time's cycle. Harvard University Press, Cambridge, MA [ Links ]
26. Gould SJ (1988) On replacing the idea of progress with an operational notion of directionality. In. Evolutionary Progress MH Nitecki (ed) pp. 319-338. University of Chicago Press, Chicago [ Links ]
27. Gould SJ (2002) The structure of evolutionary theory. The Belknap Press of Harvard University Press, Cambridge, MA [ Links ]
28. Halliwell JJ, Pérez-Mercader J, Zurek WH (1996) Physical origins of time asymmetry. Cambridge University Press, Cambridge [ Links ]
29. Hartz GA (1988) Space and time in the Leibnizian metaphysics. Noûs 22:493-519 [ Links ]
30. Hawking S (1996) A brief history of time. Bantan Books, New York [ Links ]
31. Hull DL (1975) Are species really individuals? System Zool 25:174-191 [ Links ]
32. Kant I (1804) Opus postumum. Edited by E. Förster (1993). II fascicle, VI Cambridge University Press, Cambridge [ Links ]
33. Lenski RE, Ofria C, Collier TC, Adam C (1999) Genome complexity, robustness and genetic interactions in digital organisms. Nature 400:661-664 [ Links ]
34. Lloyd-Morgan C (1926) Emergent evolution, Williams and Norgate, London [ Links ]
35. Luisi PL (1999) The interplay of cyclic and linear time in the biological world. In: Tempos in science and nature, C Rossi, S Bastanioni, A Donati, N Marchettini (eds). Ann NY Acad Sci 879:98-109 [ Links ]
36. Mackey MC (1992) Time's arrow: The origins of thermodynamic behaviour. Springer-Verlag, New York [ Links ]
37. Mcshea DW (1991) Complexity and evolution: what everybody knows. Biol & Philos 6:303-324 [ Links ]
38. Matsuno K (1989) Protobiology: physical bases of biology. CRC Press, Boca Raton, FL [ Links ]
39. Maynard-Smith J (1970) Time in the evolutionary process. Studium Generale 23:266-272 [ Links ]
40. Mayr E (1996) What is a species, and what is not? Philos Sci 63:262-277 [ Links ]
41. Nanay B (2002) The return of the replicator: what is philosophically significant in a general account of replication and selection? Biol & Philos 17:109-121 [ Links ]
42. Oliver A, Canton R, Campos P, Baquero F, Blazquez J (2000) High frequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lung infection. Science 288:1251-1253 [ Links ]
43. Peres A (1995) Quantum theory: Concepts and methods. p 406. Kluwer Academic Publishers, Dordrech [ Links ]
44. Prigogine I, Nicolis G (1977) Self-organization in non-equilibrium systems; from dissipative structures to order to fluctuations. Wiley, New York [ Links ]
45. Prigogine I (1988) La nascita del tempo. Edizioni Theoria s.r.l. Roma-Napoles [ Links ]
46. Ruse M (1998) Progress. In: The philosophy of biology, DL Hull, M Ruse (eds), pp. 589-591. Oxford University Press [ Links ]
47. Schrödinger E (1944) What is life? Cambridge University Press, Cambridge [ Links ]
48. Simpson GG (1961) Principles of animal taxonomy. Columbia University Press, New York [ Links ]
49. Spencer H (1857) Progress: its law and cause. Westminster Review 9:445-485 [ Links ]
50. Spencer H (1890) First principles, 5th edn. William and Norgate, London [ Links ]
51. Szathmary E, Maynard-Smith J (1997) From replicators to reproducers: the first major transition in life. J Theor Biol 187:555-571 [ Links ]
52. Varela FJ (1979) Principles of biological autonomy. North Holland, New York [ Links ]
53. Waddington CH (1969) Paradigm for an evolutionary process. In: Towards a theoretical biology, CH Waddington (ed), pp 106-128. Aldine Pub., Chicago [ Links ]
54. Wheeler JA (1979) Frontiers of time. North-Holland, Amsterdam [ Links ]
55. Wilson EO (1990) Success and dominance in ecosystems: the case of social insects. Ecology Institute, Oldenorf/Luhe, Germany [ Links ]
56. Yedid G, Bell G (2002) Macroevolution simulated with autonomously replicating computer programs. Nature 420:810-812 [ Links ]

