










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkArchivos de Zootecnia
versão On-line ISSN 1885-4494versão impressa ISSN 0004-0592
Arch. zootec. vol.60 no.232 Córdoba Dez. 2011
https://dx.doi.org/10.4321/S0004-05922011000400031
Atividades enzimáticasde girinos de rã-touro submetidos a rações com níveis de proteína
Enzyme activities of tadpoles of bullfrogs fed diets containing different levels of crude protein
Seixas Filho, J.T. de1,2, Oliveira, M.G.A.3A, Navarro, R.D.5, Garcia, S.L.R.4, Moura,G. de S.3B e Ribeiro Filho, O.P.3C
1Centro Universitário Augusto Motta. UNISUAM. Laboratorio de Pesquisa Biologia. Rio de Janeiro-RJ. Brasil. seixasfilho@unisuam.edu.br
2Fundação Instituto da Pesca do Estado do Rio de Janeiro. FIPERJ. Fonseca. Niterói. Brasil. jseixas@fiperj.rj.gov.br
3Universidade Federal de Viçosa. UFV. Viçosa, MG. Brasil. Amalmeida@ufv.br; Bmouraguilherme@yahoo.com.br; Coribeiro@ufv.br
4Faculdade de Viçosa. FDV. Viçosa. MG. Brasil. garciaslr@yahoo.com.br
5Faculdade de Agronomia e Medicina Veterinária. Universidade de Brasília (UnB). Brasília. Brasil. *navarrounb@gmail.com
RESUMO
Este estudo objetivou determinar a atividades das enzimas digestivas de girinos de rã-touro, alimentados com diferentes rações durante 55 dias, na densidade de um girino por litro, em caixas com 30 l de água em condições laboratoriais. Aos 18 dias os níveis de proteína influenciaram a atividade da amilase de forma quadrática até o valor estimado 32% PB. A mesma tendência foi observada para a atividade específica da amilase que influenciou até o nível de 28% de PB e da atividade da tripsina que influenciou até o nível de 29% de PB. Não foi observada diferença significativa para lípase e atividade específica da lípase. Aos 26 dias a atividade enzimática da amilase e atividade específica da amilase e da Tripsina diminuiu com aumento da porcentagem de proteína. A enzima lipase atividade específica da lípase aumentaram de forma linear ao aumento dos níveis de proteína. Aos 35 dias de experimento a atividade da amilase foi estatisticamente significativa para o tratamento com 28% de PB. A atividade específica da amilase e a atividade da tripsina diminuíram com aumento da proteína na ração. Já a atividade da lípase aos 35 e 55 dias de experimento aumentou significativamente com aumento da proteína na ração. Aos 55 dias de experimento o aumento da proteína na ração influenciou de forma linear a atividade da tripsina. A atividade da lipase e atividade específica da lipase aumentaram com a elevação do nível de proteína bruta na ração.
Palavras chave: Morfofisiologia. Nutrição animal. Ranicultura.
SUMMARY
This study was aimed to determine the digestive enzyme activity in bullfrog tadpoles, fed with commercial ration during 55 days, at the density of one tadpole per liter in 30 l boxes. After 18 days the levels of protein influenced the activity of amylase quadratically to the estimated 32% CP. The same trend was observed for the specific activity of amylase influenced by the level of 28% crude protein and the activity of trypsin that influenced by the level of 29% CP. There was no significant difference in lipase activity and specific lipase. At 26 days of the amylase enzyme activity and specific activity of amylase and trypsin decreased with increasing the percentage of protein. The enzyme lipase and lipase specific activity increased linearly to increased levels of protein. At 35 days of experiment amylase activity was statistically significant for treatment with 28% CP. The specific activity of amylase and trypsin activity decreased with increased protein in the diet. The activity of lipase at 35 and 55 days of the experiment increased significantly with increased protein in the diet. At 55 days of the experiment increased protein in the diet influenced in a linear activity of trypsin. The lipase activity and specific activity of lipase increased with increasing levels of dietary crude protein.
Key words: Morphophysiology. Animal nutrition. Tadpole production.
Introdução
Os girinos representam à fase inicial de desenvolvimento dos anfíbios e seu ciclo de vida pode ser dividido em três fases: embrionária, larval e metamorfose. A fase embrionária constitui o período da fecundação e o desenvolvimento dentro do ovo, a fase larval inicia-se com a eclosão do ovo e todo o período de desenvolvimento do girino, enquanto que a metamorfose se caracteriza pelo momento em que ocorrem mudanças do girino para a fase de adulto (Altig e McDiarmid, 1999).
A fase de criação de girinos um ponto de estrangulamento da ranicultura, devido às alterações na continuidade da produção, por doenças e mortalidades em massa, ocasionadas por procedimentos zootécnicos executados de forma empírica, principalmente a administração de alimentos inadequados, uma vez que pouco se conhece sobre as exigências nutricionais das rãs, as quais proporcionariam melhores desempenhos aos animais (Seixas-Filho, et al., 1998a, e b).
A nutrição de girinos e rãs teve como principal conquista, na década de 80, a aceitação de ração pelos animais. O desenvolvimento eficiente e saudável dos animais passa obrigatoriamente pelo fornecimento de uma dieta que satisfaz as necessidades básicas de crescimento, contendo concentrações próximas do ideal e seus diversos componentes, aliados a tecnologia de preparação. A estocagem, a concentração de macronutriente e micronutriente, a biodisponibilidade dos nutrientes, exemplos de parâmetros que interferem no desenvolvimento do animal (Navarro et al., 2007).
Os girinos se alimentam raspando com a boca, materiais aderidos ao substrato e liberando na água alimentos que posteriormente são filtrados pelas brânquias. (Kenny, 1969; Duellman e Trueb, 1986; Altig e McDiarmid, 1999).
Para se definir a formulação de rações compatíveis com as exigências nutricionais dos girinos da rã-touro, nas diferentes fases do seu desenvolvimento, é preciso conhecer a fisiologia da digestão deste animal. A eficiência de utilização dos alimentos está diretamente relacionada ao processo digestivo, no qual as enzimas têm papel fundamental.
Informações a respeito do estudo da atividade enzimática em anfíbios (Etkin, 1968, Leone et al., 1976, Hourdry et al., 1996, Bragaet al., 2004, 2006; Oliveira-Bahia, 2007, Seixas et al., 2008a e b) são escassas na literatura e existe a falta de padronização de metodologias para fornecer subsídios satisfatórios aos nutricionistas.
A importância da fase de girino para a ranicultura reside no fato que após a metamorfose existirão animais em condições compatíveis com os índices zootécnicos desejados para alta produtividade. De modo geral a criação de girinos é em ambiente aberto, onde ocorre grande produção de plâncton na água. Ao contrário dos ensaios experimentais que ocorre em ambientes fechados com total controle do ambiente.
A avaliação de rações é fundamental para que os nutricionistas possam definir um padrão alimentar, servindo de base para sistema de alimentação artificial. Nesse sentido, objetivou-se avaliar a atividades enzimáticas em quimo de girinos de rã-touro submetidos a rações contendo diferentes níveis de proteína.
Material e métodos
O experimento foi conduzido no Laboratório de Pesquisas Biológicas do Grupo AQÜISUAM da UNISUAM, Rio de Janeiro -RJ e no Laboratório de Enzimologia do Instituto de Biotecnologia Aplicada à Agropecuária- BIOAGRO da Universidade Federal de Viçosa-MG.
Durante o período experimental de 55 dias, foram utilizados 120 girinos de rãtouro (Lithobathes catesbeianus Shaw, 1802) com 15 dias, no estádio 25 de Gosner (1960), provenientes de uma mesma desova.
Os quatro módulos experimentais foram dispostos lado a lado sobre uma bancada. Os girinos foram distribuídos em caixas de polietileno (62,5 x 40 x 16 cm), com capacidade para 40 litros, de coloração branca, as quais receberam 30 l de água, acondicionando 30 animais com peso e comprimento médio inicial de, respectivamente, g e mm, resultando em uma densidade de um girino por litro.
A renovação da água nas caixas foi de 200% do volume a cada 24 horas, com abastecimento individual e mantida a uma temperatura constante de 25,0±1,0oC. O nível de água no interior das caixas foi mantido por meio de dispositivo tipo joelho adaptado na lateral das caixas, onde a água foi escoada por meio de canaletas de PVC acondicionadas lateralmente às bancadas e conectadas ao esgoto.
Cada unidade experimental recebeu aeração constante, por meio de soprador e mangueira plástica 3/16', provida de pedra porosa em sua extremidade e regulada por registro de mesmo calibre.
A ração comercial utilizada na alimentação dos girinos (tabela I) foi administrada na granulometria de 0,5mm (Seixas-Filho et al., 1998a), na proporção 10% do peso vivo distribuídas uma vez ao dia, às 12 horas, conforme Seixas-Filho et al. (1998 a e b).
A limpeza das caixas foi realizada diariamente, pela manhã (8:00h), por sifonagem de fundo, retirando-se as fezes e os restos alimentares. Diariamente, pela manhã e pela tarde, foram tomadas as temperaturas do ar e da água por meio de termômetro de coluna de mercúrio, 0 a 60oC. O controle da amônia e do pH foi efetuado diariamente, por meio colorimetria, com auxílio de kit comercial.
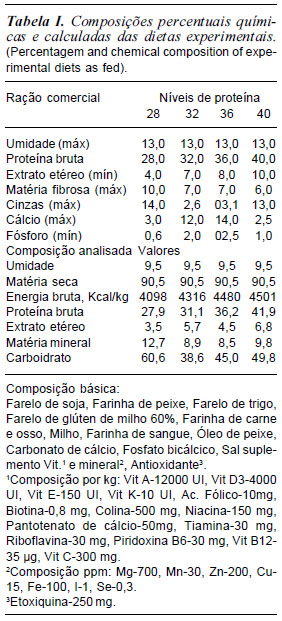
Para a realização das análises das atividades das enzimas digestivas nos quimos dos girinos, procedeu-se, no Laboratório de Pesquisas Biológicas do Grupo AQÜISUAM da UNISUAM, ao isolamento do tubo digestório dos animais e dos segmentos analisados (figura 1 - A, B, C, D, E, F).
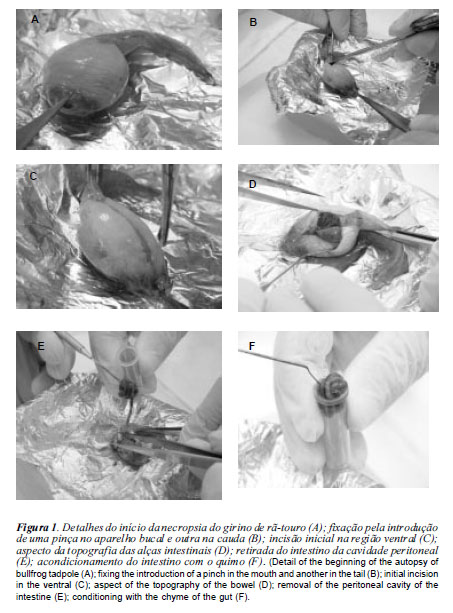
Após a verificação do estágio do girino, correspondendo as mudanças morfológicas, segundo Gosner (1960), foram coletados quatro exemplares de cada tratamento, aos 18, 26, 35 e 55 dias do experimento. Para tanto, os animais tiveram seu metabolismo reduzido por meio de imersão em banho de gelo a -4oC e anestesiados com mentol a 1% adicionado à água, em seguida, contidos fisiologicamente e submetidos a necropsia para retirada do intestino.
A necropsia dos girinos da rã-touro foi realizada em cama de gelo (figura 1A). O início da necropsia consistiu na fixação do animal com auxílio de uma pinça introduzida no interior do aparelho bucal do animal e outra no início da inserção da cauda (figura 1 B) para que houvesse resistência do tecido ä incisão do bisturi.
Os animais sofreram incisão longitudinal ventral (figura 1C) e o isolamento do tubo digestório foi feito após ligaduras duplas na porção cranial do esôfago e na porção caudal do reto, anteriormente ao ânus, para que não houvesse extravasamento do quimo.
Após a incisão visualizou-se o intestino (figura 1D). Com auxílio da espátula oblíqua, retirou-se o intestino da cavidade peritoneal, isolando-o dos outros órgãos (figura 1E), sendo acondicionado em frascos de polietileno e armazenado em freezer a -8oC até o procedimento das análises de atividade enzimática.
As análises das atividades enzimáticas foram realizadas no Laboratório de Enzimologia, Bioquímica de Proteínas e Peptídeos do Instituto de Biotecnologia Aplicada à Agropecuária (BIOAGRO) da UFV.
Para as análises de amilase, lípase e tripsina utilizaram-se 0,5 g de cada amostra congelada e 1,5 ml de tampão (Tris- HCl 0,1M), para descongelamento e maceração. Em seguida, esse material foi centrifugado a 15 000 rpm por 15 minutos a 4oC e o sobrenadante utilizado para as determinações das atividades, em duplicata.
A determinação da atividade de amilase baseou-se na hidrólise do amido pela amilase, com liberação de moléculas de glicose e dextrina, determinada em espectrofotômetro óptico, em um comprimento de onda de 660nm, utilizando-se o kit de amilase colorimétrica da Bioclin, segundo Caraway (1959). A atividade específica de amilase foi obtida pela divisão do valor da atividade de amilase pelo valor da concentração de proteína da respectiva amostra.
As atividades de lipase foram determinadas utilizando-se o kit BIOCLIN com metodologia modificada de Cherry e Crandall (1932), que consiste na verificação da atuação da lipase do quimo sobre um éster de glicerol, o qual libera um cromogênio que é quantitativamente determinado em 410 nm. Os valores são expressos em Unidades Internacionais (UI).
As atividades de tripsina foram determinadas pelo método de Erlanger et al. (1961), utilizando-se N- benzoil-D, L-arginina pnitroanilida (D, L-BApNA) como substrato.
A concentração de proteína, para todas as amostras das diferentes enzimas, foi medida na absorvância de 280 nm e os. ácidos nucléicos a 260 nm, segundo Warburg e Christin (1941).
Resultado e discussão
A temperatura do ar local apresentou média das mínimas e das máximas de 25 e 30oC, respectivamente. A média da temperatura da água observada variou de 25,0 ±1,0oC. O pH manteve-se na faixa de 7,5±0,5 e a amônia total foi de 0 à 0,25, que manteve-se dentro do desejado, sendo semelhantes aos das condições dos trabalhos com girino de Seixas-Filho et al. (1998b).
Aos 18 dias de experimento os níveis de proteína influenciaram a atividade da amilase de forma quadrática até o valor estimado 32% PB (tabela II), correspondendo ao 26o estágio larval segundo (Gosner, 1960). A mesma tendência foi observada para a atividade específica da amilase que influenciou até o nível de 28% de PB (tabela II). Estes resultados sugerem, ainda, que existe maior capacidade dos girinos em digerir alimentos à base de carboidratos em detrimento aos protéicos, sendo este fato acentuado na fase inicial da alimentação exógena deste anuro. A atividade da tripsina influenciou até o nível de 29 % de PB. Já a atividade específica da tripsina foi observada efeito linear conforme a equação:
y= 1,127292 -0,021458x, R2= 79%
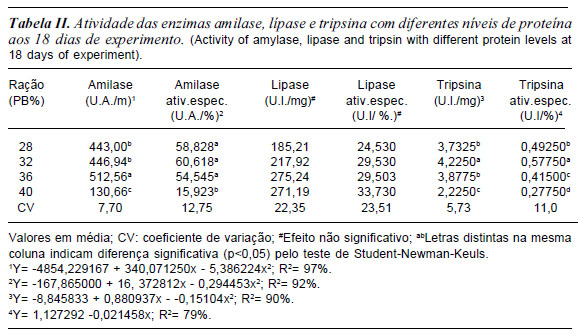
À medida que aumentava a concentração de proteína na ração, o efeito contrário foi observado para a atividade da tripsina e a atividade específica da tripsina. Entretanto não foi observada diferença significativa para lípase e atividade específica da lípase.
As enzimas pancreáticas mais encontradas durante o desenvolvimento dos girinos são tripsinas, amilases e lípases (Hourdry et al., 1996). A amilase possui maior secreção durante a fase inicial do desenvolvimento, havendo, no entanto, decréscimo da mesma no final da metamorfose. Isto ocorre em decorrência da dieta dos girinos que consomem quantidades significativas de carboidratos quando comparados com a fase adulta (carnívoro) (Leone et al., 1976).
Estudos enzimáticos mostram que existe uma estreita relação entre as alterações morfológicas do pâncreas e o perfil das enzimas de amilase e lípase. Leone et al. (1976) analisando o perfil da atividade enzimática da amilase e lípase dos girinos de Xenopus laevis, observaram que estas enzimas possuíam um pico de atividade entre os estágios 29 e 38, o qual correspondia ao maior nível de organização ultraestrutural das células acinares.
No presente estudo aos 26 dias a atividade enzimática da amilase e atividade específica da amilase diminuiu com aumento da porcentagem de proteína (tabela III). O fato da ração de 28% de proteína possuir maior quantidade de carboidratos estimulou essa maior atividade para enzima amilase (tabela III). A enzima tripsina e a atividade específica da tripsina apresentaram diminuição de atividade à medida que aumentava a proteína na ração. Estes resultados mostram que os girinos nesta fase podem ser alimentados com ração entre 28 e 32% de PB.
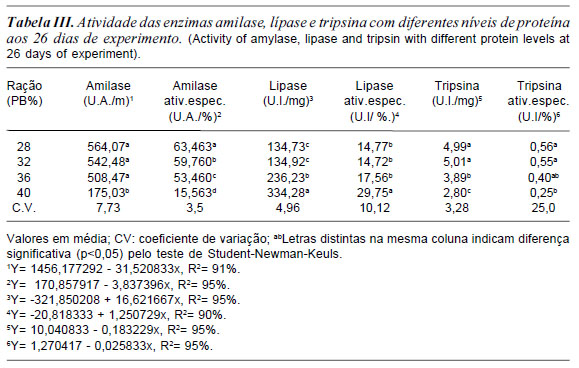
A enzima lípase e atividade específica da lípase aumentaram de forma linear ao aumento dos níveis de proteína (tabela III). Este resultado mostrou uma diminuição de 18% da atividade da lípase em comparação aos 18 dias de experimento (tabela II e III). Outro autor como Leone et al. (1976) monitoraram a atividade da lípase durante o desenvolvimento dos girinos de Xenopus laevis e registraram pico de atividade no inicio da metamorfose. Alguns estudos como de Oliveira-Bahia (2007) e Seixas Filho et al. (2008a) demonstram que o pâncreas de girino aos 21 dias após a fecundação já apresenta início da formação dos ácinos serosos pancreáticos.
O intestino é o órgão do sistema digestório que mais sofre modificação durante o processo de metamorfose. Nos estágios iniciais do desenvolvimento dos girinos, o intestino é caracterizado por um epitélio simples colunar bastante desenvolvido, denominado tecido primário, seguido por uma fina camada de tecido conjuntivo e muscular (Bonneville 1963; Marshall e Dixon, 1978; Ishizuya-Oka e Shimozawa, 1987; Hourdry et al., 1996).
Aos 35 e 55 dias de experimentos a atividade da amilase foi maior e significativa para o tratamento com 28% de PB (tabela IV e V). O fato é que a maior quantidade carboidrato estimulou essa maior atividade da enzima. No estudo de Leone et al., 1976 observaram que a amilase possui maior secreção durante a fase inicial do desenvolvimento, havendo, no entanto, decréscimo da mesma no final da metamorfose.
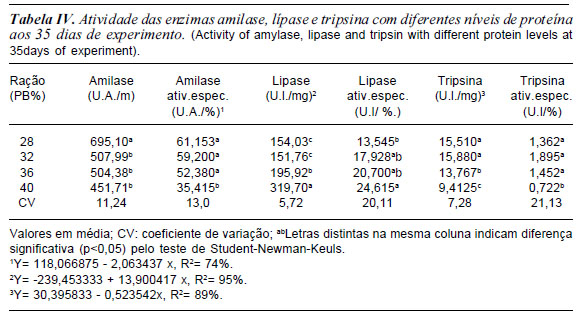
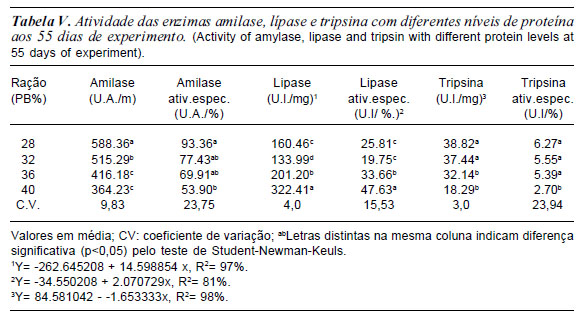
Assim como, o aumento da capacidade hidrolítica da amilase, observado no 55o dia, estão de acordo com Etkin (1968) quando afirmou que nos estágios de 36 aos 40 (fase pró-metamorfose) ocorre o maior crescimento corpóreo dos girinos, levando a um maior consumo de nutrientes e, conseqüentemente, um aporte maior de enzimas se faz necessário. Resultados semelhantes foram obtidos por Oliveira-Bahia (2007) que relatou elevado aumento na atividade de amilase, do 36o ao 40o estágio, no fígado e pâncreas de girinos de Lithobates cates-beianus, o que corresponde no presente trabalho, à última quinzena experimental.
Já a atividade da lípase aos 35 e 55 dias de experimento aumentou significativamente com aumento da proteína na ração (tabelas IV e V). Provavelmente a maior quantidade de extrato etéreo da ração em torno de 4,45 e 6,79% respectivamente, estimulou a enzima lípase, esse fato pode estar relacionado com a maior quantidade de extrato etéreo na composição do alimento, associado com a funcionalidade do pâncreas do girino neste período experimental. Podendo-se inferir que isto ocorreu pela melhor funcionalidade do pâncreas, e pelo requerimento maior de energia para o desenvolvimento dos membros posteriores nesta idade, concordando com Oliveira-Bahia (2007). Seixas Filho et al. (2008a e b) observaram em girinos a presença de células hepáticas, com arranjo glandular mais compacto e formação dos ácinos pancreáticos, o que sugere sua melhor digestão de lipídios. A atividade da lípase e atividade específica da lípase aumentou com a elevação do nível de proteína bruta na ração.
Estes resultados demonstram que, para as três enzimas em estudo, ocorreu um significativo aumento da capacidade de ação sobre o quimo dos girinos após o 260 dia de experimentação corroborando Leone et al. (1976) e Seixas Filhoet al. (2008a) em relação ao completo amadurecimento das estruturas responsáveis pela síntese e secreção das enzimas, correspondendo, neste trabalho, ao período do 15o ao 30o dia, após o início da alimentação exógena. Oliveira-Bahia, 2007, observou que amilases quanto à tripsina também apresentaram um aumento crescente durante o desenvolvimento dos girinos, ocorrendo um elevado aumento nos estágios de 36 aos 40 de Gosner (1960). Os resultados obtidos no presente estudo são basicamente semelhantes com as alterações encontradas no sistema digestório das espécies Xenopus laevis e Rana temporária (Leone et al., 1976, Ortiz De Zarete et al., 1991).
É necessário o aprimoramento das técnicas de formulação de ração para girinos para que se obtenha o maior aproveitamento do alimento oferecido, visando maiores produtividade. São recomendados também estudos simulando condições comerciais de cultivo, onde existe grande produção de plâncton no ambiente de criação, o que deve influenciar diretamente nos resultados de produção. Essas informações poderão nortear novos experimentos, melhorar as condições de cultivo, valorizar a atividade no sentido econômico, assim como fornecer embasamento a nutrição e manejo destes animais em cativeiro.
Conclusões
Em condições laboratoriais, principalmente para tripsina, entre as três enzimas em estudo, ocorreu significativo aumento da capacidade de ação sobre o quimo dos girinos após o 26o dia experimental. Na fase inicial foram verificadas maiores atividades da amilase em relação lípase e tripsina, mostrando uma boa capacidade dos girinos em digerir carboidratos, provindos da alimentação exógena. Ocorreu relação inversa da atividade de tripsina e de amilase no quimo do girino com o aumento do nível de proteína bruta na ração, para todas as fases estudadas.
Este estudo mostra a complexidade da relação entre o alimento e as enzimas, com boas respostas dos animais ao uso de rações de 28 e 32% de PB, contribuindo para o aprimoramento da alimentação artificial de rãs em cativeiro, e demonstrando a importância de novos estudos dentro do tema.
Bibliografía
Altig, R. and McDiarmid, R.W. 1999. Body plan: developmental and morphology. In: McDiarmid, R.W. and R. Altig (Eds.). Tadpoles: The biology of anuran larvae. Chicago Press. Chicago. pp. 24-51. [ Links ]
Albinati, R.C.B., Lima, S.L., Tafuri, M.L. e Donzele, J.L. 2000. Digestibilidade aparente de dois alimentos protéicos e três energéticos para girinos de rã-touro (Rana catesbeiana Shaw, 1802). Rev. Bras. Zootecn., 29: 2151-2156. [ Links ]
Albinati, R.C.B., Lima, S.L. e Donzele, J.L. 2001. Níveis de energia digestível na ração de girinos de rã-touro. Rev. Bras. Saúde Prod. Anim., 2: 48-52. [ Links ]
Bonneville, M.A. 1963. Fine structural changes in the intestinal epithelium of the bullfrog during metamorphosis. J. Cell Biol., 18: 579-597. [ Links ]
Braga, L.G.T., Oliveira, M.G.A. e Lima, W.C.F. 2004. Atividade da tripsina em rã-touro na fase pós-metamórfica. Rev. Bras. Zootecn., 33: 821-827. [ Links ]
Braga, L.G.T., Oliveira, M.G.A., Lima, W.C. e Euclydes, R.F. 2006. Atividade enzimática da lipase em rã-touro na fase pós-metamórfica. Scient. Agríc., 63: 821-827. [ Links ]
Caraway, W.T. 1959. A stable starch substrate for the determination of amylase in serum and other body fluids. Amer. J. Clin. Pathol., 32: 97-99. [ Links ]
Cherry, I.S. and Crandall Jr., L.A. 1932. The specificity of pancreatic lipase: Its appearance in the blood after pancreatic injury. Am. J. Physiol., 100: 266-73. [ Links ]
Duellman, W.E. and Trueb, L. 1968. Biology of Amphibians. McGraw-Hill. New York. pp. 508-509. [ Links ]
Etkin, W. 1968. Hormonal control of amphibian metamorphosis. In: W. Etkin and L.T. Gilbert (Eds.). Metamorphosis. A problem in development biology. Appleton Century Crofts. New York. pp. 313-348. [ Links ]
Gosner, K.L. 1960. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16: 183-190. [ Links ]
Hourdry, J., L'Hermite, A. and Ferrand, R. 1996. Changes in the digestive tract and feeding behavior of anuran amphibians during metamorphosis. Physiol. Zool., 69: 219-251. [ Links ]
Ishizuya-Oka, A. and Shimozawa, A. 1987. Development of the connective tissue in the digestive tract of the larval and metamorphosing Xenopus laevis. Anatom. Anzeiger. 164: 81-93. [ Links ]
Kenny, J.L. 1969. Feeding mechanisms in anuran larvae. J. Zool., 69: 213-251. [ Links ]
Leone, F., Lambert-Gardini, S., Sartori, C. and Scarpin, S. 1976. Ultra-structural analysis of some functional aspects of Xenopus laevis pancreas during development and metamorphosis. J. Embryol. Experim. Morphol., 36: 711-724. [ Links ]
Marshall, J.A. and Dixon, K.E. 1978. Cell specialization in the epithelium of the small intestine of feeding Xenopus laevis tadpoles. J. Anatom., 126: 133-144. [ Links ]
Navarro, R.D., Lanna, E.A.T., Donzele, J.L., Matta, S.L.P e Souza, M.A. 2007. Níveis de energia digestível da dieta sobre o desempenho de piauçu (Leporinus macrocephalus) em fase pós-larval. Acta Sci. Anim. Sci., 29: 109-114. [ Links ]
Oliveira-Bahia, V.R.L. 2007. Morfologia e enzimologia do sistema digestório dos girinos de rã-touro (Rana catesbeiana) durante o desenvolvimento e metamorfose. Tese Doutorado. Universidade Estadual Paulista. Jaboticabal. 148 pp. [ Links ]
Ortiz de Zarete, A., Villaro, A.C., Etayo, J.C., Diaz de Rada, O., Montuenga, L.M., Sesma, P. and Vazquez, J.J. 1991. Developmental of the endocrine pancreas during larval phases of Rana temporaria: An immunocytochemical and ultrastructural study. Cell Tissue Res., 264: 139-150. [ Links ]
Seixas-Filho, J.T., Mello, S.C.R.P., Veiga, R.C.A., Miranda, R.G.B. e Santos, C.A.N. 1998a. Efeito da granulometria da ração sobre o desempenho de girinos de Rana catesbeiana. Rev. Bras. Zootecn., 27: 224-230. [ Links ]
Seixas-Filho, J.T., Mello, S.C.R.P., Silva, J.M.F., Tomas, J.E. e Melo, C.M.S. 1998b. Efeito dos níveis de energia e proteína bruta no desempenho de girinos (Rana catesbeiana Shaw, 1802). Rev. Bras. Zootecn., 27: 664-669. [ Links ]
Seixas Filho, J.T, Gomes, L.H., Aguiar, D.V.C, Hipólito, M., Martins, A.M.C.R.P. da F. e Chaves, A.C.P. 2008a. Avaliação histológica do intestino médio, do fígado e do pâncreas de girinos de rãtouro alimentados com rações comerciais formuladas com três níveis de proteína bruta. Rev. Bras. Zootecn., 37: 2090-2096. [ Links ]
Seixas Filho, J.T, Hipólito, M., Carvalho, V.F., Martins, A.M.C.R. da F., Silva, L.N. e Castagna, A.A. 2008b. Alterações histopatológicas em girinos de rã-touro alimentados com rações comerciais de diferentes níveis protéicos. Rev. Bras. Zootecn., 37: 2085-2089. [ Links ]
Warburg, O. and Christin, W. 1941. Isohering und kristallisation dês görungs ferments enolase. Biochem. Zool., 310: 384-421. [ Links ]
Recibido: 22-2-10
Aceptado: 8-2-11.

