










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkArchivos de Zootecnia
versión On-line ISSN 1885-4494versión impresa ISSN 0004-0592
Arch. zootec. vol.60 no.229 Córdoba mar. 2011
https://dx.doi.org/10.4321/S0004-05922011000100012
Desarrollo cardiaco embrionario en broilers en relación con enfriamiento y altitud
Poultry's embrionary heart development in relation whit cooling and altitude
Camacho-Escobar, M.A.1* y Suárez-Oporta, M.E.2
1Instituto de Industrias. Universidad del Mar. Campus Puerto Escondido. Ciudad Universitaria. Puerto Escondido. CP 71980. Mixtepec. Juquila. Oaxaca. México. *marcama@zicatela.umar.mx
2Colegio de Postgraduados. Campus Tabasco. Calle Castaños no 303 Fraccionamiento Los Reyes Loma Alta. H. Cárdenas. Tabasco. CP 86570. México.
RESUMEN
Se estudió la relación entre enfriamiento y altitud durante la incubación y el desarrollo cardiaco embrionario. Se incubaron huevos de pollo de engorda a 2250 m de altitud, en tres tratamientos: no enfriados (NE), enfriado a temperatura ambiente 6 o 12 h (E6 y E12) los días 10, 12, 14, 16 y 18; se compararon con embriones incubados en condiciones comerciales a 900 m (COM). El día 19 de incubación se sacrificaron 24 embriones por cada tratamiento, para determinar daño cardiaco ocasionado por frío e hipoxia. Los resultados indican que no hubo diferencia en peso del embrión, el peso ajustado del corazón fue mayor en el tratamiento COM (p<0,01); el largo y ancho del corazón se afectaron (p<0,01) por el enfriamiento presentando una relación directa con el tiempo de enfriamiento. Las correlaciones entre variables fueron altas. La duración del enfriamiento prolongó la incubación. El enfriamiento y la altitud de incubación afectaron el desarrollo cardiaco, posiblemente debido a las condiciones de hipoxia de los embriones estudiados durante la incubación.
Palabras clave: Crecimiento compensatorio. Desarrollo embrionario. Hipertermia. Hipertrofia ventricular. Hipoxia.
SUMMARY
The relationship between cool and altitude during incubation in embrionary heart development was studied. Chicks' eggs were hatched at 2250 m altitude, in three treatments: no cooled (NE), cooled at room temperature for 6 or 12 h in 10, 12, 14,16, and 18 incubation day (E6 and E12), there were compared with embryos hatched in commercial conditions at 900 m (COM). The 19 hatch day 24 embryos were culled for each treatment to determine heart demage by cool and hypoxia. The results showed no differences in embryo weight, the adjusted heart weight was mayor in COM treatment (p<0.01); heart length and width were affected (p<0.01) by the cool and showed directly relation with the cooling time. The correlations between variables were high; the cool time extended the hatch time. The cool and incubation altitude affected the heart development, maybe by the hypoxic conditions in the embryos studied during the hatch.
Key words: Compensatory growth. Embrionary development. Heat stress. Heart hypertrophy. Hypoxic.
Introducción
Las modernas líneas comerciales de pollo de engorda tienen tasa metabólica más acelerada durante todo el desarrollo embrionario y las primeras semanas de vida del pollo, que otras líneas genéticas de aves (Hoer, 1988), debido a ello, la necesidad de oxígeno en todo su organismo es mayor (Odom et al., 1992). Mediante técnicas de selección genética se ha logrado incrementar el tamaño del huevo y consecuentemente, el tamaño del embrión; sin embargo, el número y tamaño de los poros en el cascarón no han sido considerados como variable respuesta en los programas de mejoramiento genético (Black y Snyder 1980; Decuypere y Michels, 1992). En la actualidad se incuban huevos con embriones de mayor velocidad de crecimiento que los incubados hace 100 años (Christensen et al., 2000); pero proporcionalmente con menor superficie de intercambio gaseoso (Burton y Tullet, 1983), esta situación provoca que el embrión tenga insuficiencia en la capacidad de intercambio gaseoso para cubrir sus demandas durante el último tercio del desarrollo (Christensen y McCorkle, 1982). Ello provoca condiciones de hipoxia para el embrión (Maxwell et al., 1987), similares a las presentadas en huevos incubados a grandes alturas (Thompsom, 1952).
Aunado a dicha hipoxia, es posible que en las actuales condiciones de incubación se presenten condiciones de hipertemia durante el desarrollo embrionario provocado por el inadecuado intercambio gaseoso entre el interior y exterior del huevo que dificulta la disipación del calor generado por el metabolismo embrionario. La temperatura interna del huevo se incrementa aproximadamente 0,2oC por día a partir del día 11 de incubación, alcanzando hasta 40oC al final de la misma (Romijn y Lokhorst, 1956; Tullet, 1990); en ocasiones, el gradiente de temperatura que existe entre el embrión dentro del huevo y el ambiente de la incubadora, no permite la disipación del calor metabólico (Van Brecht et al., 2005), esta situación puede provocar hipertermia embrionaria (Romijn y Lokhorst, 1956).
Al enfriar el huevo durante la última etapa de la incubación, se reduce la actividad metabólica del embrión, la cual antes de nacer es un tercio de la tasa metabólica de un ave adulta con igual masa corporal (Hoyt et al., 1975), proporcionalmente su capacidad de producir calor es menor (Lancaster y Jones, 1988; Subba y Glick, 1977; Tullet, 1990). Se ha reportado que los tratamientos de enfriamiento del embrión, disminuyen la generación de calor metabólico, su consumo de O2 (Black y Snyder, 1980) y la cantidad de hormonas tiroideas circulantes (Chris-tensen et al., 2002).
Existen evidencias de que la susceptibilidad del pollo de engorda a la ascitis está relacionada con factores predisponentes originados durante el desarrollo embrionario, como la hipoxia (Maxwell et al., 1987; Odom et al., 1991; Odom et al., 1992; Tona et al., 2005) y la tasa metabólica acelerada que tiene el embrión (Hoer, 1988). La hipoxia embrionaria es un factor predisponente al desarrollo de la hipertensión pulmonar crónica adaptativa, condición común en pollos ascíticos, provocando estrés cardiaco antes o durante el nacimiento (Arce et al., 1987; Odom et al., 1992; Odom, 1993). Al disminuirse experimentalmente la conductancia del cascarón, el embrión desarrolla adaptaciones fisiopatológicas similares a las de aves con ascitis, y los pollos obtenidos de dichos huevos son más propensos a desarrollar la enfermedad (Maxwell et al., 1987; Odom, 1993). Estudios en pollitos de un día de edad incubados en condiciones comerciales, indican la presencia de hipertrofia ventricular adaptativa, lesión típica en pollos ascíticos, con incidencia de 10 a 50% (Odom et al., 1991).
Para comprobar si es posible reducir la tasa metabólica del embrión mediante enfriamiento del huevo en la segunda mitad del desarrollo embrionario, y con ello reducir daños cardiacos en el embrión, el objetivo del presente trabajo fue determinar el efecto del enfriamiento y la altitud sobre el nivel del mar de la incubadora, en la presentación de hipertrofia ventricular derecha en embriones de pollo de engorda.
Material y métodos
Se incubaron huevos de pollo de engorda comercial, en las instalaciones del Programa de Ganadería del Colegio de Post-graduados, ubicadas en Montecillo, municipio de Texcoco, Estado de México a una altitud de 2250 m, y se compararon con embriones procedentes de una incubadora comercial ubicada en Jiutepec, Estado de Morelos, a 900 metros sobre el nivel del mar.
Durante los meses de septiembre a octubre, se incubaron 900 huevos provenientes de reproductoras Ross x Ross de 32 semanas de edad; se utilizó una máquina incubadora marca Iamex con capacidad para 2300 huevos y con volteo automático cada hora. Al arribo de los huevos fértiles al cuarto de incubación en Montecillo, procedentes de la incubadora comercial; se identificaron individualmente con numeración continua en ambos extremos. Se pesaron con una balanza sensibilidad de 0,001 g.
Posteriormente fueron colocados en charolas de incubación, distribuídas aleatoriamente dentro de la incubadora con temperatura interna constante de 37,5oC (bulbo seco) y 30oC (bulbo húmedo). Cuando todos los huevos estuvieron dentro de la incubadora se cerró y ese momento fue considerado como el inicio de la incubación.
Se aplicaron tres tratamientos con 12 repeticiones de 25 huevos; los tratamientos aplicados fueron: incubación continua sin enfriar (NE); enfriamiento durante 6 horas continuas los días 10, 12, 14, 16 y 18 de incubación (E6) y enfriamiento durante 12 horas continuas los mismos días que el tratamiento anterior (E12).
El enfriamiento se hizo a temperatura ambiente, sacando directamente las charolas con los huevos de la incubadora y colocándolas sobre una mesa en el cuarto de incubación; transcurrido el tiempo de enfriamiento, las charolas fueron reintroducidas a la incubadora.
La temperatura dentro del cuarto de incubación durante el enfriamiento fue de 18oC (± 2oC). El día 19 de incubación, los huevos se sacaron de la incubadora y mediante ovoscopia se determinó la infertilidad o mortalidad embrionaria. Los huevos con embriones vivos fueron puestos en canastas nacedoras con divisiones individuales para mantener la identificación del pollito al eclosionar, y retornados a la incubadora. La humedad relativa de la misma se aumentó a 80% y la temperatura se disminuyó a 37,2oC. Cuando se cumplieron 21 días de incubación se comenzó a pesar y marcar los pollitos eclosionados con intervalos de 6 h, al cumplirse el día 22 de incubación se apagó la incubadora y en ese momento se consideró terminada la incubación. Las variables consideradas para los pollitos al eclosionar fueron: peso del huevo, peso del pollito al eclosionar y peso al final de la incubación.
Al momento de pasar los huevos a las canastas nacedoras, se seleccionaron al azar 24 huevos con embrión vivo, dos huevos de cada repetición de los tres tratamientos, se colocaron en una bolsa plástica, la cual se llenó de CO2 con el fin de sacrificar inmediatamente a los embriones. Se realizaron mediciones del peso, largo y ancho del corazón con la finalidad de determinar la presencia de hipoxia debido a la altitud durante la incubación. Para efectos de comparación, se obtuvieron de la incubadora comercial 25 huevos con embriones viables, del mismo lote de los huevos incubados en las instalaciones del Colegio de Postgraduados y con igual tiempo de incubación.
De cada embrión se obtuvo el corazón, a cada uno de ellos, se le retiró el atrio, los vasos mayores y el pericardio para su medición. En embriones sacrificados se midieron las siguientes variables: peso del embrión, largo y ancho del corazón (Julian et al., 1987). Las medidas del corazón se hicieron mediante un calibrador con sensibilidad de 1 mm.
Los tratamientos se distribuyeron en un diseño completamente al azar y se analizaron mediante el paquete estadístico SAS (SAS Institute, 1988). Se utilizó el procedimiento General Linear Model (GLM) y para determinar la diferencia entre tratamientos se usaron contrastes ortogonales. Se usó el peso del embrión como covariable para ajustar el peso, largo y ancho del corazón. El cálculo de las correlaciones entre variables continuas se hizo con el procedimiento Correlation (CORR).
Se consideraron las diferencias entre tratamientos a nivel de p<0,05 y p<0,01.
Resultados
Bajo las condiciones de incubación con altitud, los tratamientos de enfriamiento (E6 y E12) provocaron que el peso del embrión sacrificado al día 19 de incubación, fuera menor (p<0,01) respecto al tratamiento no enfriado (NE); similarmente, el tratamiento E6 presentó mayor peso que el tratamiento (E12), dicha diferencia fue altamente significativa (p< 0,01). Este comportamiento se debe al retraso de crecimiento que se provocó en el desarrollo embrionario durante los tratamientos de enfriamiento, retraso que fue directamente proporcional al de la duración del enfriamiento; sin embargo, debido a que el peso de los embriones incubados en condiciones comerciales (COM) fue igual que el tratamiento NE, al realizar el contraste ortogonal, no presenta diferencia estadística (tabla I).

Al ajustar el peso del corazón con respecto al peso del embrión, para evitar el efecto de comparar embriones y corazones con diferente peso, los embriones incubados en condiciones comerciales (COM), presentaron significativamente mayor peso del corazón (p<0,01), respecto a los corazones de embriones incubados a mayor altitud, con o sin enfriamiento (NE, E6 y E12), sin que hubiera diferencia entre los tratamientos incubados a mayor altitud; esta situación indica que la mayor altitud de incubación, afecta el desarrollo cardiaco del embrión de pollo provocando órganos más ligeros (tabla I).
De manera similar el corazón de los embriones incubados en condiciones comerciales (COM) presentaron corazones de menor tamaño en sus dimensiones de largo y ancho, respecto a los corazones de embriones incubados a mayor altitud; dichas diferencias fueron altamente significativas (p<0,01), lo cual puede indicar que aún durante el desarrollo embrionario, se requiere de ajustes en la presión sanguínea para oxigenar los tejidos, provocando con ello distensión del músculo cardiaco (tabla I).
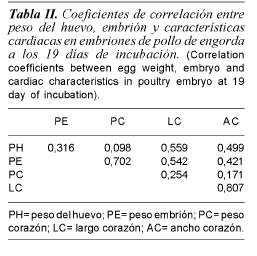
Respecto a las características del corazón, únicamente el largo y ancho muestran mediana correlación (tabla II), del mismo modo que el peso del corazón con el peso del embrión.
Los datos de la tabla III indican que no hubo variación significativa en el peso de los huevos y que los tratamientos de enfriamiento no afectaron estadísticamente el peso del pollito al momento de eclosionar; sin embargo, muestra diferencias altamente significativas (p<0,01) en el peso del pollito al final de la incubación, lo cual indica que los pollitos que no tuvieron retraso en su desarrollo embrionario, presentaron mayor deshidratación por permanecer más tiempo en la nacedora.

Discusión
Se ha reportado que las condiciones de incubación a 1500 m sobre el nivel del mar provocan hipoxia moderada, y que la incubación en altitud de 3800 m tiene efectos de hipoxia severa en el embrión (Black y Snyder, 1980); sin embargo, esta situación no afecta el crecimiento del embrión, debido a que los pollitos incubados a esas alturas son de igual peso que los incubados a nivel del mar; similares resultados se obtuvieron entre el tratamiento COM y el E12 (tabla I). Las condiciones de hipoxia están relacionadas con la presentación de hipertrofia cardiaca (Hoer, 1988), similar a la hipertrofia ventricular derecha, representativa en condiciones de ascítis en pollos de engorda (Wood, 1991). En el presente trabajo, los embriones con enfriamiento prolongado (E6 y E12) presentaron corazones más largos y más anchos (p<0,01) que los del tratamiento COM, indicando que las condiciones de altitud y enfriamiento provocaron hipertrofia cardiaca en dichos embriones, no así en el tratamiento que tuvo únicamente condiciones de altitud (NE); esta hipertrofia cardiaca puede ser debido al incremento en el gasto cardiaco del embrión, por ajustes metabólicos compensatorios para oxigenarse (Decuypere, 1979). Los resultados indican que existe correlación entre las dimensiones cardiacas del embrión, por lo cual se puede inferir que al realizar ajustes en la presión sanguínea, se presenta una deformación en el ventrículo derecho que también provocará incremento en la longitud cardiaca (tabla II).
En general, las aves son más tolerantes a la hipoxia que los mamíferos, ello se debe a adaptaciones fisiológicas en su sistema respiratorio, reajustes del transporte de O2 en la sangre, modificaciones a nivel vascular cerebral (Faraci, 1991), y mayor afinidad de la hemoglobina al O2 (Black y Snyder, 1980). Los animales, cuando sufren hipoxia, reducen su tasa metabólica como respuesta adaptativa para reducir la demanda de O2 y proteger a órganos sensibles a su deficiencia, como cerebro y corazón (Wood, 1991); sin embargo, algunos autores señalan que esta baja en la tasa metabólica puede representar simplemente la incapacidad del individuo para obtener el O2 requerido (Carey et al., 1982). Es posible, que en los últimos días de incubación exista un equilibrio entre la tolerancia a la hipoxia y la capacidad de reducir la intensidad respiratoria por unidad de masa corporal (Decuypere y Michels, 1992).
Los embriones sometidos a hipoxia severa, presentan compensaciones adaptativas como reajuste vascular del flujo sanguíneo, principalmente en las regiones pobremente oxigenadas de la membrana corioalantóidea, e incrementos en el gasto cardiaco (Odom et al., 1992). En condiciones de hipoxia, el gasto cardiaco puede alterarse (Wood, 1991), lo cual depende de los reflejos simpáticos o de alteraciones en la contractibilidad cardiaca (Tazawa et al., 1991). Se han reportado diferencias significativas en el peso de corazones de pollos recién eclosionados provenientes de huevos sometidos a condiciones de hipoxia moderada o severa (Black y Snyder, 1980), lo anterior se confirmó en el presente trabajo al encontrarse diferencias altamente significativas (p<0,01) en el peso de corazones de embriones incubados a de 2250 m de altitud (tabla I) respecto al peso de los corazones de embriones incubados a 900 m. Al incubar huevos en condiciones de menor O2 disponible, el corazón de los embriones tiene que realizar mayor gasto cardiaco, provocándose con ello, aumento en el tamaño como resultado de una hipertrofia (Odom et al., 1992), lo que es congruente con los datos obtenidos en el presente estudio.
Los tratamientos de enfriamiento no afectaron significativamente el peso del pollito al nacer (tabla III), a pesar que se han reportado mayores pesos al eclosionar de huevos sometidos a tratamientos de enfriamiento durante la incubación, lo que se atribuye a la presentación de crecimiento compensatorio embrionario al terminar el tratamiento de enfriamiento (Geers et al., 1983); en este trabajo dicha condición no se presentó, posiblemente porque el último día de enfriamiento fue muy cercano al momento de la eclosión, y no se permitió la manifestación del crecimiento compensatorio en el embrión.
Al enfriar huevos a temperatura de 22oC durante 6, 12 o 18 h en el día 16 de incubación, no se reportan diferencias significativas en el peso al nacimiento entre los tratamientos de enfriamiento y el testigo (Sarpong y Reinhart, 1985), similares resultados se reportan considerando que el embrión compensa el crecimiento (Yalcin y Siegel, 2003); sin embargo, en otro experimento, al someter a tratamiento de enfriamiento huevos de pollo de engorda a temperatura de 10oC, durante 30 minutos en el día 12 o 16 de incubación, los pollitos obtuvieron mayores pesos iniciales (p< 0,05) que los pollitos del tratamiento no enfriado (Al-Hassani, 1993), pero no se reporta si existieron diferencias en el peso de los pollitos al momento de la eclosión. Esta información difiere con otro reporte que indica menor calidad, peso y vitalidad del pollito, al enfriar embriones a menos de 21oC (Lancaster y Jones, 1988).
El peso de los pollitos al final de la incubación tuvo una relación inversa con el tiempo de enfriamiento (p<0,01), por ello en los tratamientos sometidos a enfriamiento más prolongado, los pollitos eclosionaron más tarde respecto a los pollitos provenientes de tratamientos con menor tiempo de enfriamiento o sin enfriar (tabla III). Los pollitos del tratamiento NE eclosionaron durante las primeras horas del día 21 de incubación, los pollitos del tratamiento E6 a partir de las primeras 12 h de dicho día y los del tratamiento E12 durante las últimas horas del mismo día. Esto explica, porqué los pollitos provenientes del tratamiento E1, presentaron el menor peso al final de la incubación, debido a que se deshidrataron al permanecer más tiempo en las canastas nacedoras sin acceso a agua y con temperatura de 37,2 oC.
El tiempo de incubación se aumentó en los huevos de tratamientos enfriados, se ha reportado que al enfriar el huevo fértil durante la incubación el desarrollo embrionario se retrasa y el tiempo de incubación se alarga un período de tiempo igual al del enfriamiento (Booth y Rahan, 1991); sin embargo, otros investigadores señalan que el retraso en tiempo es menor, debido a que la temperatura interna del huevo no cambia bruscamente durante los períodos de enfriamiento (Suárez, 1993) y el embrión es capaz de compensar su crecimiento (Yalcin y Siegel, 2003).
Se concluye, que existen indicios de que la incubación a 2250 m de altitud, provee al embrión las condiciones de hipoxia suficientes para afectar las características del corazón de los embriones de pollo de engorda, haciéndolos más ligeros, largos y anchos al compararlos con los corazones de embriones incubados a 900 m de altitud del tratamiento comercial. Esto puede indicar, que las condiciones de hipoxia provocadas por la altitud a la que se encuentre la incubadora, sí afectan el desarrollo cardiaco del embrión, sin embargo, es necesario realizar mayor investigación sobre el tema.
Agradecimientos
Al Consejo Nacional de Ciencia y Tecnología por su apoyo económico para la realización de la presente investigación.
Bibliografía
Al-Hassani, D. 1993. Production and biochemical studies on chick embryos under cold stress. Indian J. Anim. Sci., 63: 46-49. [ Links ]
Arce, J., López, C. y Vásquez, C.G. 1987. Análisis de la incidencia del síndrome ascítico en el Valle de México. Técnica Pecuaria en México, 25: 338-346. [ Links ]
Black, C.P. and Snyder, G.K. 1980. Oxygen transport in the avian egg at high altitude. Am. Zool., 20: 461-468. [ Links ]
Booth, D.T. and Rahan, H. 1991. Effects of periodic egg cooling during incubation. In: Tullet, S.G. (comp.). Avian incubation, 1st ed. Butterworth-Heinemann. England. [ Links ]
Burton, F.G. and Tullet, S.G. 1983. A comparison of the effects of eggshell porosity on the respiration and growth of domestic fowl duck and turkey embryos. Comp. Biochem. Phys. A., 75: 167-174. [ Links ]
Carey, C., Thompsom, E.L., Vleck, C.M. and James, F.C. 1982. Avian reproduction over an altitudinal gradient: incubation period, hatching mass, and embryonic oxygen consumption. The Auk, 99: 710-718. [ Links ]
Christensen, V.L., Davis, G.S. and Nestor, K.E. 2002. Environmental incubation factors influence embryonic thyroid hormones. Poultry Sci., 81: 442-450. [ Links ]
Christensen, V.L., Grimes, J.L., Donaldson, W.E. and Lerner, S. 2000. Correlation of body weight with hatchling blood glucose concentration and its relationship to embryonic survival. Poultry Sci., 79: 1817-1822. [ Links ]
Christensen, V.L. and McCorkle, F.M. 1982. Turkey egg weight losses and embryonic mortality during incubation. Poultry Sci., 61: 1209-1213. [ Links ]
Decuypere, E. and Michels, H. 1992. Incubation temperature as a management tool: a review. World Poultry Sci. J., 48: 28-38. [ Links ]
Decuypere, E. 1979. Effect of incubation temperature patterns on morphological, physiological and reproduction criteria in Rhode Island red birds. Agricultura, 27: 65-68. [ Links ]
Faraci, F.M. 1991. Adaptations to hypoxia in birds: how to fly high. Annu. Rev. Physiol., 53: 59-70. [ Links ]
Geers, R., Michels, H., Nackaerts, G. and Konings, F. 1983. Metabolism and growth of chickens before and after hatch in relation to incubation temperatures. Poultry Sci., 62: 1869-1875. [ Links ]
Hoer, F.J. 1988. Patogenesis of ascites. Poultry Digest., 1: 8-12. [ Links ]
Hoyt, D.F., Vleck, D. and Vleck, C.M. 1975. Metabolism of avian embryos: ontogeny and temperature effects in the ostrich. Condor, 80: 265-271. [ Links ]
Julian, R.J., Friars, G.W., French, H. and Quinton, M. 1987. The relationship of right ventricular hypertrophy, right ventricular failure, and ascities to weight gain broiler and roaster chickens. Avian Dis., 31: 130-135. [ Links ]
Lancaster, F.M. and Jones, D.R. 1988. Cooling of broiler hatching eggs during incubation. Brit. Poultry Sci., 29: 597-604. [ Links ]
Maxwell, M.H., Tullet, S.G. and Burton, F.G. 1987. Haematology and morphological changes in young broiler chicks with experimentally induced hypoxia. Res. Vet. Sci., 43: 331-338. [ Links ]
Odom, T.W., Hargis, B.M., López, C.C., Arce, J.M., Ono, Y. and Avila, G.E. 1991. Use of electrocardiographic analysis for investigation of ascites syndrome in broiler chickens. Avian Dis., 35: 738-744. [ Links ]
Odom, T.W., Rosembaum, L.M., Stolz, J.L. and Jeong, D. 1992. Experimental reduction of egg shell conductance during incubation. III. Attenuation of the effect on early ascites syndrome mortality by removal of the egg shell cuticle. Poultry Sci., (Abs): 71: 1. [ Links ]
Odom, T.W., Rosenbaum, L.M. and Hargis, B.M. 1992. Evaluation of electrocardiographic analysis of young broiler chickens as a predictive index for susceptibility to ascites syndrome. Avian Dis., 36: 78-83. [ Links ]
Odom, T.W. 1993. La relación entre la genética, la incubación y el ambiente después del nacimiento con el desarrollo del síndrome ascítico en el pollo de engorda. En: XVIII Convención Nacional ANECA. Cancún, Q.R. pp. 168-176. [ Links ]
Romijn, C. and Lokhorst, W. 1956. The caloric equilibrium of the chicken embryo. Poultry Sci., 35: 829-834. [ Links ]
Sarpong, S. and Reinhart, B.S. 1985. Broiler hatching stress and subsequent growout performance. Poultry Sci., 64: 232-234. [ Links ]
SAS Institute. 1988. SAS/SAT User's Guide (Release 6.03). SAS Inst. Inc. Cary NC. USA. [ Links ]
Suárez, M.E. 1993. Factors affecting incubation time and chick quality [Doctoral dissertation]. University of Florida. Gainesville. Florida. USA. [ Links ]
Subba Rao, D.S.V. and Glick, B. 1977. Effect of cold exposure on the inmune response of chickens. Poultry Sci., 56: 992-996. [ Links ]
Tazawa, H., Hiraguchi, T., Takura, T., Yamaguchi, S., Yamada, M. and Yufu, Y. 1991. The cardiogenic ballistogram of embryos - non-invasive determination of the heart rate. In: Tullet, S.G. (comp.) Avian incubation, 1st ed. Butterworth-Heinemann. England. [ Links ]
Thompsom, R.L. 1952. Incubation at high altitudes. The effects of wind, barometric pressure and humidity on foetal mortality in the hens egg. Poultry Sci., 31: 497-508. [ Links ]
Tona, K., Kemps, B., Bruggeman, V., Bamelis, F., De Smith, L., Onaghbesan, O., De Baerde-maeker, J. and Decuypere, E. 2005. Comparison of three lines of broiler breeders differing in ascites susceptibility or growth rate. 1. Relationship between acoustic resonance data and embryonic or hatching parameters. Poultry Sci., 84: 1439-1445. [ Links ]
Tullet, S.G. 1990. Science and art of incubation. Poultry Sci., 69: 1-5. [ Links ]
Van Brecht, A., Hens, H., Lemaire, J.L., Aerts, J.M., Degraeve, P. and Berchmans, D. 2005. Quantification of the heat exchange of chicken eggs. Poultry Sci., 84: 353-361. [ Links ]
Wood, S.C. 1991. Interactions between hypoxia and hypothermia. Annu. Rev. Physiol., 53: 71-85. [ Links ]
Yalcin, S. and Siegel, P.B. 2003. Exposure to cold or heat during incubation on developmental stability or broiler embryos. Poultry Sci., 82: 1388-1392. [ Links ]
Recibido: 15-7-08.
Aceptado: 22-7-09.

