










 Servicios personalizados
Servicios personalizados
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
En los pacientes con enfermedad renal crónica (ERC) los mecanismos de regulación de la homeostasis del calcio y del fósforo se hallan comprometidos, lo que da lugar a lo que se conoce como “alteraciones minerales y óseas de la enfermedad renal-crónica”, más conocidas por su acrónimo en inglés, CKD-MBD (Chronic Kidney Disease-Mineral and Bone Disorders), que incluye la calcificación vascular (CV) (1).
La CV se asocia a un incremento de la mortalidad en la población general (2) y principalmente en pacientes con ERC (3,4), en los que es más prevalente que en individuos de la misma edad con función renal preservada (5,6).
La elevada morbimortalidad de la ERC se ha atribuido a factores tradicionales y no tradicionales de riesgo. Al igual que en la población general, los factores de riesgo tradicionales (tabaquismo, hipertensión, diabetes y sexo masculino) son responsables en gran medida de la progresión de la CV. Sin embargo, estos factores no son suficientes para explicar la elevada prevalencia de las complicaciones cardiovasculares en pacientes con ERC (7). Entre los factores no tradicionales, la hiperfosfatemia es uno de los más estudiados relacionados con el aumento de CV y de mortalidad en los pacientes con ERC (8,9).
La CV se produce a través de un proceso complejo, activo y regulado en el que intervienen diferentes mecanismos (10-12). Hasta la fecha se han publicado diversos estudios que han permitido avanzar en el conocimiento de los mecanismos subyacentes al proceso de CV (7,10). Sin embargo, la imposibilidad de una detección precoz de la CV ha impulsado la investigación sobre nuevos posibles biomarcadores que puedan utilizarse para el diagnóstico temprano de la CV y que podrían mejorar sus estrategias preventivas y pronósticas. La espectrometría de masas aplicada a estudios de proteómica permite la caracterización del perfil proteico (proteoma) de una muestra biológica de forma rápida en un momento determinado. Por tanto, el objetivo fundamental de este trabajo fue estudiar la expresión diferencial de proteínas asociadas al proceso de CV mediante técnicas de proteómica utilizando un modelo de CV in vitro con células de músculo liso vascular (CMLV).
MATERIALES Y MÉTODOS
CULTIVO CELULAR Y CONDICIONES DE CALCIFICACIÓN
Se utilizó como CMLV la línea celular inmortal A7r5 de aorta de rata (ATCC CRL-1444™). Las células se cultivaron en DMEM (1,8 mM de calcio [Ca] y 1 mM de fosfato [P]; Lonza) suplementado con suero fetal bovino (SFB) (10 %), glutamina (1 %) (Biochrom), aminoácidos no esenciales (ANE) (1 %), penicilina y estreptomicina (1 %) (Biochrom).
Las células A7r5 se sembraron en 20 placas de 15 cm de diámetro (152 cm2 de superficie). Para el análisis del contenido en calcio se usaron placas de seis pocillos (9,6 cm2 de superficie). Cuando alcanzaron el 60-70 % de confluencia, se añadieron las condiciones experimentales. Para los experimentos de calcificación se utilizó el medio de cultivo DMEM-F12 para maximizar las diferencias entre las condiciones no calcificantes y calcificantes. Medio no calcificante (control): DMEM-F12 (1 mM de Ca y P) suplementado con BSA (0,1 %), penicilina y estreptomicina (1 %). Medio calcificante: al medio no calcificante se le añadió un estímulo calcificante para que las células A7r5 cambiaran su fenotipo a tipo osteoblasto; en este caso se suplementó con Ca y P hasta una concentración final de 2 y 3 mM, respectivamente. En nuestras condiciones experimentales, concentraciones superiores a 2 mM de calcio y a 3 mM de fosfato producen una precipitación espontánea de Ca3(PO4).
Ambos estudios se realizaron en paralelo y el medio de cultivo se renovó cada dos días.
CONTENIDO EN CALCIO
El grado de mineralización se valoró mediante tinción con rojo de alizarina a los 6 días de cultivo en las condiciones calcificantes y no calcificantes. Brevemente, las células se lavaron con phosfate buffer saline (PBS), se fijaron con formaldehido (10 % en PBS) a 4 °C durante 45 minutos y, tras un lavado con agua desionizada, se tiñeron con rojo de alizarina al 2 % (pH del 4,2) durante 5 minutos.
ESTUDIO DE PROTEÓMICA
Tras 6 días de cultivo y una vez comprobada la presencia de calcificación, las células se recogieron en 20 mL de PBS y se juntaron para obtener 4 muestras cultivadas en medio no calcificante (C1, C2, C3 y C4) y cuatro muestras del cultivo en MC (P1, P2, P3 y P4). En ambos grupos las muestras se solubilizaron mediante sonicación en tampón de lisis (7M urea, 2M tiourea, 4 % CHAPS y 30 mM Tris). La concentración total de proteínas se medió utilizando el método Bradford (13) (Bio-Rad, Hercules, CA, EE. UU.).
Siguiendo las recomendaciones del fabricante se añadieron 400 pmol de CyDyeTM DIGE FluorDyes (GE Healthcare, Uppsala, Suecia) por 50 µg de proteína (los fluorocromos Cy3 y Cy5 se usaron para la tinción de las muestras y Cy2, para el estándar interno, resultado de la mezcla de la misma cantidad de proteína de las muestras cultivadas en condiciones no calcificantes y calcificantes). Las muestras se tiñeron por pares (condiciones no calcificantes y calcificantes) para evitar diferencias en el marcaje.
La separación de los extractos de proteínas marcadas se llevó a cabo en geles bidimensionales DIGE (2D Fluorescence Difference Gel Electrophoresis), según las recomendaciones del fabricante (GE Healthcare, Uppsala, Suecia). Primera dimensión: las muestras se cargaron en tiras IPG (24 mm; pH de 3-10 NL) (GE Healthcare). Una vez realizada la primera dimensión, las tiras IPG se incubaron en un tampón de equilibrado (50 mM Tris-HCl, pH de 8,8, 6M urea, 30 % glicerol, 2 % SDS y trazas de azul de bromofenol) que contenía 0,5 % ditiotreitol (DTT) durante 15 minutos y a continuación, en el mismo tampón, con 4,5 % iodoacetamida otros 15 minutos. Para la segunda dimensión, las tiras se cargaron en geles de poliacrilamida (12,5 %) (Ettan DALT six system, GE Healthcare, Uppsala, Suecia) y se corrieron (2W/gel) durante 4 horas y 30 minutos hasta que el azul de bromofenol alcanzó el extremo del gel. Después, los geles 2D fueron escaneados (EttanDigerImager, GE Healthcare, Uppsala, Suecia) a 100 µm de resolución con λex/λem de 488/520, 532/580 y 633/670 nm para Cy2, Cy3, y Cy5, respectivamente.
Se realizaron tres experimentos independientes para cada condición. El análisis de las imágenes se llevó a cabo con el programa informático DeCyder2D v7.0 (GE Healthcare, Uppsala, Suecia). Para la selección de spots, la cuantificación del volumen y la normalización de las muestras en el gel se usó el módulo de análisis diferencial (DIA o differential in-gel analysis). El módulo de análisis de variación biológica (BVA o biological variation analysis) se usó para comparar los spots de proteínas entre los distintos geles e identificar aquellos que mostraron diferencias significativas.
Se realizó un gel preparativo en poliacrilamida (12,5 %) con 500 µg de proteína (la misma cantidad de cada muestra) siguiendo el mismo procedimiento descrito anteriormente. Las proteínas se visualizaron mediante tinción con Oriole™ Fluorescent Gel Stain (Biorad, Hercules, EE. UU.); la adquisición de las imágenes se realizó mediante un documentador de geles ChemiDoc™ XRS+ (Biorad, Hercules, EE. UU.).
Los spots expresados diferencialmente se cortaron manualmente. La digestión de los spots se realizó con 12,5 ng/µl tripsina en 50 mM bicarbonato de amonio durante 12 horas a 37 °C. Para la extracción de los péptidos se utilizó una disolución de ácido fórmico (1 %) y acetonitrilo (50 %) (Sigma Aldrich, St Louis, EE. UU.).
IDENTIFICACIÓN DE PROTEÍNAS
Los extractos de proteínas se analizaron usando un sistema de nanoHPLC (Ultimate 3000, Dionex/LC Packings) con muestreador automático conectado a un espectrómetro de masas de trampa iónica (ion Q-trap) (Applied Biosystems) con fuente de nanoelectrospray (ESI). La columna cromatográfica fue C-18 de 75 µm diámetro (Dionex/LC Packings) conectada a un capilar de óxido de silicio para generar el electroespray. El volumen de inyección de la muestra utilizado fue 1 µl y el flujo de la fase móvil, 0,125 mL/min (modo split).
Los espectros de masas (MS/MS) obtenidos se analizaron con el software Analyst y se enfrentaron a la base de datos SwissProt para la identificación de las proteínas usando el motor MASCOT (MatrixScience). Los parámetros de búsqueda incluyeron una tolerancia de ± 1,2 Da y posibles modificaciones de las proteínas, como deamidación, carbametilación y oxidación de la metionina. Se consideró que una proteína estaba identificada cuando se detectaron al menos dos péptidos diferentes. La función y la localización de las proteínas identificadas se asignaron de acuerdo con la información depositada en las bases de datos PubMed y ExPasy.
En los datos suplementarios se muestra un esquema de los procesos que se realizaron en el estudio de proteómica (Fig. 1) y las imágenes representativas de los geles marcados con los fluorocromos Cy2, Cy3 y Cy5 y la imagen resultante de la unión de los tres fluorocromos (Fig. 2).
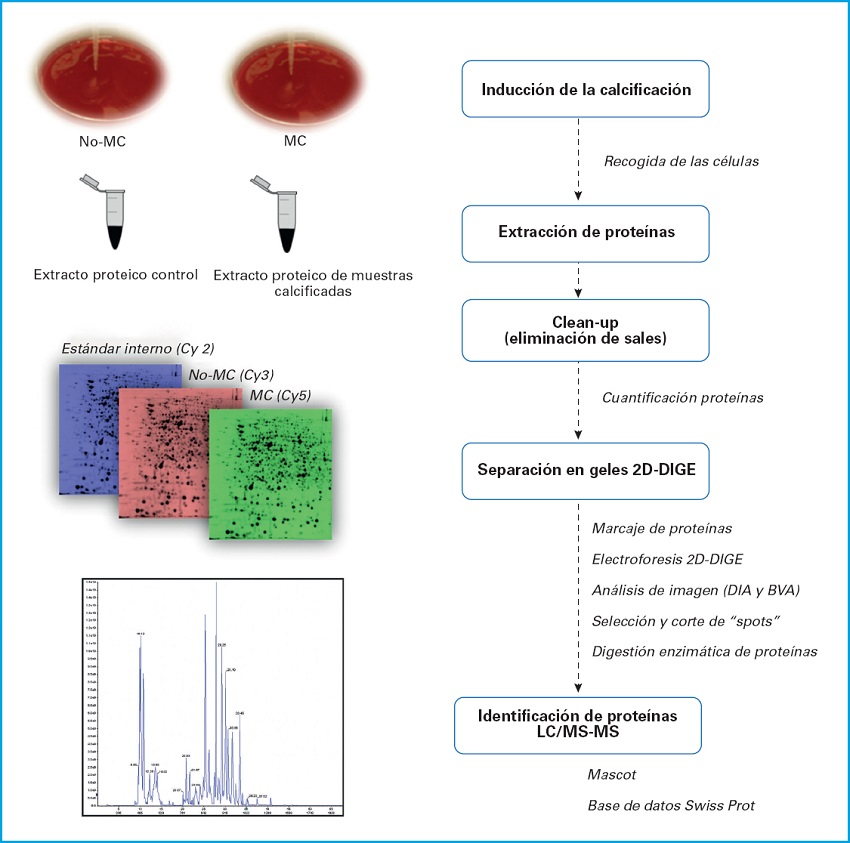

ANÁLISIS ESTADÍSTICO
El análisis de los resultados del DIGE se realizó con el programa informático DeCyder2D v7.0 (GE Healthcare, Uppsala, Suecia) mediante análisis estadístico T-Student (p < 0,05). Se seleccionaron los spots de proteínas, que mostraron cambios en la expresión por encima de un umbral de 1,5.
RESULTADOS
Las condiciones experimentales de cultivo de las CMLV se mantuvieron hasta el sexto día de tratamiento, momento en el que se observó, mediante la tinción con rojo de alizarina, un aumento en la deposición de calcio en los cultivos tratados con medio calcificante (Fig. 3).
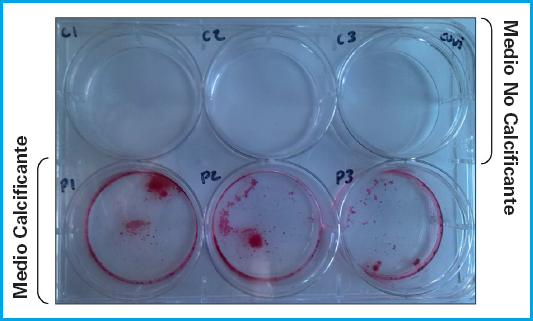
Figura 3. Tinción con rojo alizarina de las CMLV tras 6 días de cultivo en condiciones no calcificantes y calcificantes.
Para evaluar la expresión diferencial de proteínas en cultivo de CMLV tratadas con medio calcificante respecto a las células cultivadas en condiciones no calcificantes se llevó a cabo el análisis comparativo 2D- DIGE. Aproximadamente, se detectaron 1860 spots de proteínas, de los que 121 se expresaron diferencialmente (p < 0,05), y de estos, 101 spots se analizaron por LC-ESI-MS/MS (Fig. 4). Los espectros de masas obtenidos se enfrentaron a las bases de datos de modo que una puntuación individual superior a 32 obtenida en el Mascot indicó identidad u homología (p < 0,05). Se identificaron un total de 20 proteínas diferentes en 24 spots.

Figura 4. Mapa del perfil de proteínas expresadas de forma diferencial en cultivos de CMLV en condiciones calcificantes respecto a las no calcificantes. Gel preparativo. Se indica el número de spot y el símbolo MASCOT de las proteínas identificadas.
De las proteínas identificadas, cuatro aumentaron su expresión en las células cultivadas en medio calcificante respecto a las cultivadas en medio no calcificante (Tabla I). El colágeno de tipo I y la anexina A2 mostraron las mayores diferencias entre las condiciones no calcificantes y calcificantes (ratio 3,49 y 2,17, respectivamente) después de 6 días.
Tabla I. Proteínas sobreexpresadas en cultivos de CMVL A7r5 cultivadas en condiciones calcificantes respecto a las cultivadas en condiciones no calcificantes e identificadas por LC-MS/MS.
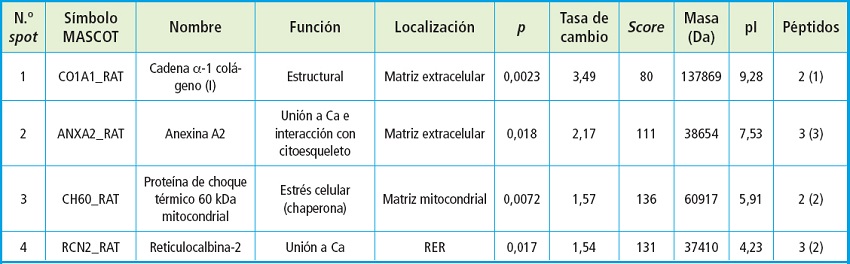
RER: retículo endoplasmático rugoso. MS Se muestra el número de spot, el símbolo del Mascot, la función y la localización de la proteína y el valor teórico de la masa (Da) y el punto isoeléctrico (pI) obtenido de las bases de datos, es decir, sin tener en cuenta posibles alteraciones o modificaciones postraslacionales. Otros datos del análisis por espectrometría de masas, como el número de péptidos identificados, también fueron incluidos (el número de péptidos únicos se indican entre paréntesis).
En la tabla II se muestran las 16 proteínas cuya expresión resultó inhibida en células cultivadas en medio calcificante respecto a las cultivadas en medio no calcificante. Las tropomiosinas (cadena beta, alfa-1 y alfa-3) fueron las proteínas que mostraron un mayor descenso cuando las CMLV se cultivaron en medio calcificante. Además, algunas de ellas se identificaron en varios spots, indicando posibles alteraciones en su estructura o modificaciones postraduccionales. Otras proteínas propias de células musculares que redujeron su expresión fueron la actina, la miosina-9, la desmina o la alfa-actina y otras proteínas del citoesqueleto, como la vimentina o la cadena alfa-1-A de la tubulina.
Tabla II. Proteínas inhibidas en cultivos de CMVL A7r5 sometida a tratamiento de calcificación respecto al control.
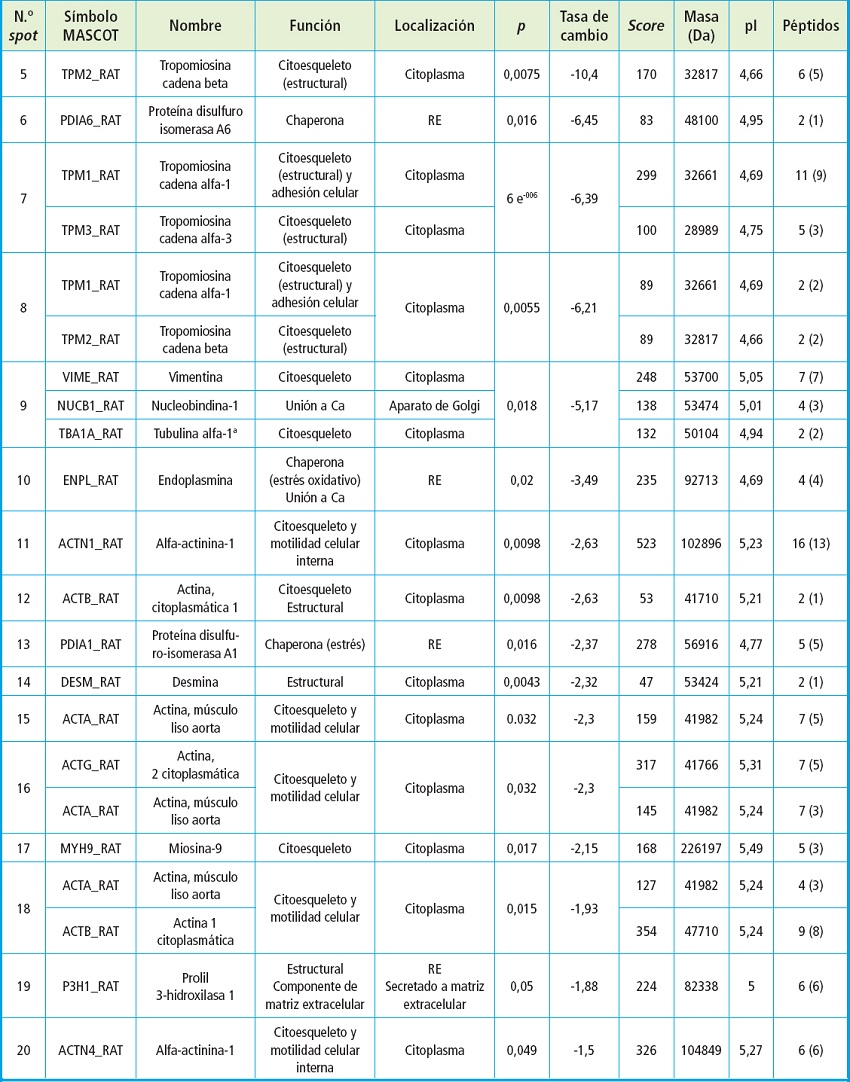
RE: retículo endoplasmático.
La proteína disulfuro isomerasa A6 y A1 y la endoplasmina fueron tres chaperonas localizadas en el retículo endoplásmico que redujeron su expresión más de seis, tres y dos veces, respectivamente, en las condiciones calcificantes respecto a las no calcificantes. También se observó una disminución en la expresión de nucleobindina-1, proteína de unión a calcio localizada en el aparato de Golgi y P3H1 que tiene función estructural y es un componente de la matriz extracelular.
DISCUSIÓN
La publicación de numerosos estudios sobre CV y ERC ha permitido conocer parcialmente el proceso de CV asociado a ERC. En este sentido, las técnicas ómicas (genómica, transcriptómica y proteómica, junto con otras técnicas de análisis) proporcionan una poderosa herramienta para obtener e integrar información biológica del proceso de CV (14). Las proteínas, principales efectores de la mayor parte de los procesos biológicos, son las moléculas más adecuadas para usarse como marcadores o dianas para el tratamiento de las enfermedades debido a una relación estrecha entre las proteínas y los fenotipos. Por este motivo los resultados obtenidos en los estudios proteómicos podrían ser más reproducibles que los de genómica y la transcriptómica (14).
Tras el análisis por espectrometría de masas de los extractos proteicos obtenidos de las CMLV tras 6 días de cultivo en medio calcificante y no calcificante se identificaron 20 proteínas con una fiabilidad elevada.
De las cuatro proteínas identificadas que vieron aumentada su expresión en medio calcificante, la que presentó mayor cambio fue la cadena alfa del colágeno de tipo I. Hay varios trabajos que relacionan el aumento de los niveles del colágeno I y la CV. El colágeno I y III son los principales componentes de la matriz extracelular cuyos niveles se mantienen relativamente estables en condiciones fisiológicas. Sin embargo, niveles elevados de fosfato y de calcio aumentan la expresión de colágeno I, lo que favorece la diferenciación de las CMLV a células de tipo osteoblasto (15). Otra proteína implicada en la biosíntesis y en la maduración del colágeno es la prolil-3-hidroxilasa (P3H1), proteoglicano con actividad enzimática necesaria para la correcta formación de las hélices de colágeno, y cuya expresión se redujo en condiciones calcificantes, coincidiendo con otros trabajos en los que también se analiza el perfil de expresión proteico en CMLV en condiciones calcificantes (16). Además, ratones deficientes en esta proteína mostraron una hipermineralización de la matriz ósea (17). Cambios en las características de la matriz extracelular y en el comportamiento entre los constituyentes de la matriz influyen no solo en las propiedades mecánicas de los tejidos conectivos, sino que contribuyen a modular el fenotipo celular, alterando la expresión de proteínas, la organización del citoesqueleto y, en consecuencia, las rutas de señalización intracelulares.
Otras proteínas del RER cuya expresión se encuentra alterada son las proteínas disulfuro isomerasa A1 y A6 (PDIA1, PDIA 6) implicadas en la formación de enlaces disulfuro, la isomerización y la reducción, actuando junto con chaperonas en la regulación de proteínas mal plegadas. La expresión de estas proteínas se encontró inhibida en condiciones calcificantes, y aunque no hay evidencias del papel de la PDIA6 en la CV, cambios en la expresión de PDIA1 se han asociado con un aumento de la CV y aórtica (18,19).
Dentro de este grupo de proteínas involucradas en el plegamiento y cuya expresión se encontró modificada en este estudio se encuentra la proteína de choque térmico de 60 kDa mitocondrial (CH60) y la endoplasmina (ENPL). La expresión de la proteína HSP 60 en la enfermedad vascular se ha correlacionado con la severidad del proceso de aterosclerosis y la respuesta antiinflamatoria (20). Varios estudios han demostrado que la expresión de HSP está estrechamente relacionada con el proceso de CV (21-23). Por otra parte, aunque no se ha descrito asociación entre la ENPL y la CV, esta proteína participa en la diferenciación normal de las células musculares cardíacas, lisas y esqueléticas (24), por lo que cambios en su expresión podrían alterar su diferenciación normal.
En el presente estudio también se observó un aumento en dos proteínas de unión a Ca+2, como la reticulocalbina-2 (Rcn-2) y la anexina A2. Las proteínas de unión a Ca (CaBP o calcium binding protein) a través de dominios específicos participan activamente en muchos procesos celulares, como la homeostasis del Ca+2 o rutas de señalización (25). La iniciación de la mineralización requiere la entrada de Ca+2 en las vesículas de la matriz. En concreto, Rcn-2 se ha descrito como un mediador de la CV que aumenta la expresión de marcadores osteogénicos y reduce la de marcadores de fenotipo contráctil en CMLV (26). Por otro lado, las anexinas son una familia de proteínas de unión a membrana dependientes de Ca. Diferentes estudios han mostrado la importancia de las anexinas en la formación de vesículas de la matriz a través de la entrada de Ca y del inicio de la mineralización (27) y las semejanzas existentes entre el proceso de formación de vesículas de matriz a partir de las membranas celulares de las CMLV y los condrocitos, incluyendo el enriquecimiento de las anexinas A2, A5 y A6 como desencadenante del proceso de mineralización (28). Otra proteína de unión a calcio, pero que en este caso se encontró inhibida, es la nucleobindina 1 (NUCB1), localizada en el aparato de Golgi, donde desempeña un importante papel en la homeostasis del Ca (29), aunque hasta el momento no hay trabajos que la asocien con la CV.
Asimismo, en concordancia con trabajos previos llevados a cabo en modelos in vivo e in vitro de CV y proteómica, los resultados de este trabajo mostraron un descenso en la expresión de proteínas estructurales y en la desregulación de las proteínas del citoesqueleto en las CMLV (16,30,31). En este grupo de proteínas, las que experimentaron mayor descenso fueron las proteínas típicamente musculares (tropomiosina, actina, desmina, miosina, etc.). Este hecho puede interpretarse como una pérdida del fenotipo muscular de las CMLV como consecuencia de la exposición a altas concentraciones de P y Ca, ya descrito en otros trabajos (32).
Uno de los problemas de la CV es que se diagnostica en etapas tardías, una vez que ya está establecida, y hasta la fecha no se han descrito terapias que se dirijan específicamente al tratamiento de la CV. Además, las técnicas de diagnóstico empleadas, principalmente basadas en técnicas de imagen, pueden estar limitadas debido a factores como el coste, la disponibilidad y la exposición a la radiación (33). Por tanto, la identificación de biomarcadores circulantes mínimamente invasivos, sensibles y específicos que puedan identificar la presencia de CV en etapas tempranas podría presentarse como una alternativa de gran interés. Las proteínas identificadas en este trabajo podrían ayudar en la búsqueda de los biomarcadores que permitan identificar a los pacientes con mayor riesgo de desarrollar CV, o incluso podrían utilizarse como dianas terapéuticas una vez validados en otros estudios.
Este tipo de estudios presenta una serie de limitaciones, ya que la identificación de proteínas está limitada por la cantidad y la calidad (poca concentración de sales, ácidos nucleicos, lípidos, etc.) del extracto proteínico obtenido, lo que depende directamente de la eficacia del método de extracción, y la preparación de las muestras, que son fundamentales para la calidad de los resultados (34). En este trabajo, además de las proteínas que se identificaron con alta fiabilidad, también se detectaron otras con una puntuación baja, lo que podría deberse a diferentes razones (35): en los spots cortados la concentración de algunas proteínas puede ser muy inferior respecto a otras mayoritarias y las modificaciones postranscripcionales o proteólisis de algunas proteínas podrían alterar la disponibilidad de los péptidos para su identificación. También hay que tener en cuenta que, al tratarse de un modelo de calcificación in vitro, no permite una comparación directa entre el estado de ERC y un control sano, ya que la ERC implica factores sistémicos y condiciones fisiológicas que no se capturan completamente en este modelo. Otra limitación a considerar es que la línea celular utilizada fue de rata, por lo que la traslación de los resultados a humanos es limitada. Por otra parte, la caracterización del perfil proteico de las células se ha llevado a cabo en un modelo de calcificación que se mantuvo durante seis días. Análisis en otros puntos del proceso contribuirían a profundizar en el conocimiento de la calcificación vascular. Es posible que en modelos en los que la calcificación pudiese mantenerse durante períodos más prolongados los niveles de algunas proteínas podrían haber sido mayores.
CONCLUSIONES
Los resultados obtenidos confirmaron un cambio en la expresión fenotípica de las células musculares, con un descenso de las proteínas típicamente musculares y del citoesqueleto. Además, fue posible identificar cambios en la expresión de proteínas que previamente no se habían relacionado con la CV y que podrían participar en dicho proceso. Teniendo en cuenta los resultados obtenidos en este trabajo, sería de gran utilidad realizar experimentos complementarios que permitiesen esclarecer el papel de estas y de otras proteínas expresadas diferencialmente en el proceso de CV. Estas proteínas podrían utilizarse como marcadores del estadio precoz de la enfermedad o como moléculas diana para el desarrollo de nuevas estrategias terapéuticas que permitan reducir la morbimortalidad de pacientes con ERC.

