










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkArchivos de Zootecnia
versão On-line ISSN 1885-4494versão impressa ISSN 0004-0592
Arch. zootec. vol.63 no.243 Córdoba Set. 2014
https://dx.doi.org/10.4321/S0004-05922014000300010
Parámetros genéticos para caracteres productivos y reproductivos en Holstein y Jersey colombiano
Genetic parameters for production and reproduction traits in Colombian Holstein and Jersey cattle
Zambrano, J.C.1*; Rincón, J.C.2A y Echeverri, J.J.2B
1Posgrado en Biotecnología. Universidad Nacional de Colombia sede Medellín. Medellín. Colombia. *jczambrano@unal.edu.co
2Departamento de Producción Animal. Universidad Nacional de Colombia Sede Medellín. Medellín. Colombia. Ajcrincon@unal.edu.co; Bjjecheve@unal.edu.co
Se agradece a la Universidad Nacional de Colombia sede Medellín, a la Cooperativa Colanta Ltda. y al Ministerio de Agricultura y Desarrollo Rural por su financiación y valioso apoyo en la realización de este trabajo.
RESUMEN
El objetivo de este estudio fue estimar parámetros genéticos y fenotípicos en características productivas y reproductivas en ganado Holstein y Jersey de Antioquia, Colombia. La información fue obtenida de 85 hatos lecheros de raza Holstein y 7 hatos de raza Jersey, ubicados en 18 municipios del departamento de Antioquia. Las características productivas evaluadas fueron: producción de leche (PL), porcentaje de proteína (PP), porcentaje de grasa (PG) y puntaje de células somáticas (SCS) y las características reproductivas evaluadas fueron: intervalo entre partos (IEP), días abiertos (DA), número de servicios por concepción (NSC) y tasa de concepción (TC). Los componentes de (co)varianza, los parámetros genéticos y fenotípicos fueron estimados empleando modelos mixtos univariados y bivariados y las soluciones de las ecuaciones de los modelos mixtos fueron obtenidas mediante el método de máxima verosimilitud restricta libre de derivadas. Las heredabilidades obtenidas para características productivas oscilaron entre 0,22 y 0,35, excepto para SCS que fueron bajas (h2<0,11) y para las características reproductivas oscilaron entre 0,03 y 0,15. Las correlaciones genéticas más altas obtenidas para Holstein fueron: rgIEPxPL= 0,77; rgIEPxPP= 0,61; rgIEPxPL= 0,77 y rgDAxSCS= 0,73 y para Jersey: rgIEPxPP = 0,56; rg|EPxSCS= 0,49; rgTCxPG= - 0,52 y rgTCxSCS= -0,72. Las correlaciones fenotípicas fueron bajas en todos los casos, variaron entre -0,17 y 0,16. Las características productivas, son mucho más heredables que las características reproductivas. No obstante, las correlaciones genéticas sugieren que es recomendable construir índices de selección a partir de estas características para obtener un mayor progreso genético pero equilibrado en sus componentes productivos y reproductivos.
Palabras clave: BLUP. Heredabilidad. Repetibilidad.
SUMMARY
The objective of this study was to estimate genetic and phenotypic parameters in milk production and reproduction traits in Holstein and Jersey from Antioquia, Colombia. The information was obtained from 85 Holstein dairy herds and 7 Jersey dairy herds. The milk production traits evaluated were: milk yield (PL), protein percentage (PP), fat percentage (PG) and somatic cell score (SCS) and the reproduction traits evaluated were: calving interval (IEP), days open (DA), number of services per conception (NSC) and conception rate (TC). The (co) variance components, genetic and phenotypic parameters were estimated by using univariate and bivariate linear mixed models and the solutions of the mixed model equations were obtained through derivative-free restricted maximum likelihood method. The heritabilities obtained for milk production traits ranged from 0.22 to 0.35, except for SCS that were low (h2<0.11) and for reproduction traits ranged between 0.03 and 0.15. The highest genetic correlations for HolStein Were: rgIEPxPL= 0,77; rgIEPxPP= 0,61; rgIEPxPL= 0,77 and rgDAxSCS= 0,73 and for Jersey were: rgIEPxPP = 0,56; rg|EPxSCS= 0,49; rgTCxPG= - 0,52 y rgTCxSCS= -0,72. Phenotypic correlations were low in all cases, ranged from -0.17 to 0.16. Milk production traits are more heritable than reproduction traits. However, genetic correlations suggest that it is favorable to construct selection indices from these traits to gain more genetic progress but balanced in their productive and reproductive components.
Key words: BLUP. Heritability. Repeatability.
Introducción
Las características reproductivas en hembras bovinas productoras de leche tienen gran importancia, ya que afectan en gran medida la rentabilidad de los hatos lecheros especializados. Estas características tienen en común que sus heredabilidades son muy bajas y como consecuencia su progreso genético es muy lento (Sbardella y Gaya, 2010 y Camargo, 2012). Las pérdidas económicas por problemas reproductivos son atribuidas principalmente a intervalo entre partos prolongados, incremento en los costos de inseminación, pocos terneros por vaca por año, incremento de descarte de animales, elevados costos de reemplazo y menor vida productiva de las vacas (Wall et al., 2003 y Camargo, 2012). A pesar de los problemas mencionados, el desempeño reproductivo de los animales no fue incluido por muchos años en programas de mejoramiento genético a nivel mundial y una de las causas fue que la selección se orientó hacia la producción lechera, con excepción de los países escandinavos, en donde en los índices de selección además de las características productivas, se incluyeron características reproductivas y de salud (Miglior et al., 2005). Sin embargo, en los últimos años, algunos países han incluido características de fertilidad en sus índices de selección y consideran que su incorporación en esquemas de mejoramiento genético en ganado de leche tiene gran importancia (Ka-darmideen etal., 2003 y Walsh et al., 2011).
Determinar cuáles características reproductivas incluir en las evaluaciones genéticas es difícil porque son características complejas, afectadas por múltiples factores y de baja heredabilidad (Jamrozik et al., 2005; Walsh et al., 2011). Como consecuencia, la selección directa para características reproductivas podría ser ineficiente, como consecuencia de la baja precisión de los valores genéticos, especialmente para vacas y toros jóvenes, ocasionando que las decisiones de selección para estas características frecuentemente involucren gran in-certidumbre (Jamrozik et al., 2005 y Sewalem et al., 2010). Por el contrario, las características productivas y de calidad de la leche tiene heredabilidades medias y altas y su selección involucra precisiones mucho más elevadas. Pero existe un antagonismo genético entre características productivas y reproductivas que perjudica la fertilidad de las vacas cuando éstas han sido seleccionadas por su alto mérito genético para producción lechera (Jamrozik et al., 2005). Sewalem et al. (2010) recomiendan una evaluación conjunta entre características reproductivas y producción de leche usando modelos multicaracterísticos para incrementar la precisión de la selección. No obstante, esto es posible solo cuando existen altas asociaciones genéticas entre las características (Henderson y Quaas, 1976; Mrode y Thompson, 2005).
En Colombia, las investigaciones relacionadas con la estimación de parámetros genéticos y evaluaciones genéticas para características productivas y principalmente para características reproductivas en ganado lechero son muy limitadas. Por lo tanto, es muy importante desarrollar estudios que involucren la estimación de parámetros genéticos y fenotípicos en condiciones propias del trópico alto colombiano que incluyan además de características productivas, características asociadas a fertilidad. El objetivo de este estudio fue estimar parámetros genéticos y fenotípicos en características productivas y características reproductivas en ganado Holstein y Jersey de Antioquia, Colombia.
Material y métodos
Esta investigación fue avalada por el Comité de Ética en Investigación de la Universidad Nacional de Colombia sede Medellín, considerando que cumple con los estándares establecidos para este tipo de investigación (Número de la carta de aprobación: CEMED-015 Mayo, 2012).
POBLACIÓN Y FUENTE DE DATOS
Se estudió la información de 85 hatos de raza Holstein y 7 de raza Jersey, ubicados en 18 municipios del Departamento de Antioquia (Colombia), que cubre zonas de bosque muy húmedo montano bajo (bmh-mb), que tiene como límites climáticos generales una temperatura aproximada entre 12 y 18 oC y un promedio anual de lluvias entre 2000 y 4000 mm. Normalmente se extienden en una faja altimétrica de 1800 a 2800 msnm. Las condiciones específicas de manejo, alimentación y sanidad fueron variables en todos los hatos, así como su topografía y ubicación geográfica. La información reproductiva y sanitaria de los hatos (fechas de parto, fechas de secado, fecha de inseminación, fechas de detección de preñez, entre otras) fue obtenida de los registros históricos, libretas y software de manejo ganadero.
Las características reproductivas que fueron evaluadas en esta investigación fueron: intervalo entre partos (IEP) medido como el número de días que transcurrieron entre dos partos consecutivos, número de servicios por concepción (NSC) medido como el número de inseminaciones que fueron llevadas a cabo hasta detectar la preñez de las hembras, días abiertos (DA) medido como los días que transcurrieron desde el parto a la concepción y la tasa de concepción (TC), la cual fue definida como el porcentaje de éxito de que las inseminaciones llevadas a cabo fueron satisfactorias, [TC= (1/NSC)*100].
Por otra parte, las características productivas evaluadas en esta investigación fueron: producción de leche (PL, medida en litros/lactancia), porcentaje de proteína (PP) porcentaje de grasa (PG) y recuento de células somáticas (RCS, media en células/mL) respectivamente. El recuento de células somáticas fue transformado a puntaje de células somáticas (SCS) usando la siguiente ecuación: SCS= [log2 (RCS/100 000)] + 3, con el fin de mejorar la normalidad de los datos, como describen Ali y Shook (1980). Toda la información fue digitalizada, analizada, y almacenada en Software Control 1, versión 1.0 (Echeverri et al., 2010), del laboratorio de mejoramiento genético animal de la Universidad Nacional de Colombia. Los datos extremos considerados fisiológicamente anormales o registrados erróneamente o que generaban sospecha de su invalidez, fueron eliminados. El número de animales con información productiva e información reproductiva y el número de registros por vaca para todas las características evaluadas en las razas Holstein y Jersey se indican en la tabla I. Los animales que fueron incluidos en la matriz de parentesco para la raza Holstein fue de 7937, de los cuales 593 fueron padres (con un promedio de 4 hijas por padre) y 755 madres. Para la raza Jersey, el número de animales incluidos en la matriz de parentesco fue de 833 de los cuales 93 fueron padres (con un promedio de 5 hijas por padre) y 108 madres.

ANÁLISIS ESTADÍSTICO
La normalidad de los datos en todas las características incluidas en esta investigación, fue evaluada mediante las pruebas de Kolmogorov-Smirnov, Cramer-von Mises y Anderson-Darling, usando el procedimiento PROC UNIVARIATE de SAS (2006). También se evaluó la independencia de los errores y la homogeneidad de varianzas; esta última fue evaluada mediante el test de Levene. Una vez validados los supuestos de normalidad, se llevó a cabo un análisis descriptivo en el cual se estimó la media, la desviación estándar y el coeficiente de variación para todas las características en la raza Holstein y en la raza Jersey.
ESTIMACIÓN DE LA HEREDABILIDAD Y LA REPETIBILIDAD
Los parámetros genéticos (heredabilidad y repetibilidad) fueron estimados en la raza Holstein y en la raza Jersey, empleando un modelo animal univariado (Mrode y Thompson, 2005), independiente para cada una de las características. La notación matricial del modelo fue:
y= Xb + Za + Wap + e
donde:
y = vector de observaciones (PL, PG, PP, SCS, DA, IEP, NSC y TC);
b= vector de efectos fijos dentro de los cuales fueron incluidos: el efecto fijo hato (96 hatos en Holstein y 7 hatos en Jersey), el efecto fijo número de parto (vacas entre 1 y 8 partos en Holstein y entre 1 y 4 partos en Jersey) y el efecto fijo del grupo contemporáneo (municipio-mes de parto-año de parto) dentro del cual fueron conformados 123 grupos en Holstein y 31 grupos en Jersey con al menos 5 registros cada uno. Dentro del vector de efectos fijos, también fueron incluidas algunas covariables como: duración de la lactancia solo para la característica PL y producción de leche solo para las características PP y PG.
Los efectos aleatorios incluidos en el modelo mixto fueron:
a= vector del efecto aleatorio genético aditivo;
ap= vector del efecto aleatorio del ambiente permanente;
e= vector del efecto aleatorio del residual;
X, W y Z= matrices de incidencia que relacionan los registros con los efectos fijos y aleatorios.
Se asume que los efectos aleatorios de: ambiente permanente, genético aditivo y del residual son distribuidos independientemente con media cero y varianzas:

considerando que:
A= matriz de relaciones genéticas aditivas y Iσ2e=R, se tiene que:

Las ecuaciones del modelo mixto para el estimador lineal insesgado (BLUE) de las funciones estimables de (b) y para el mejor predictor lineal insesgado (BLUP) de (ap) y a fueron descritas como:

donde:
I= matriz identidad;

El modelo animal univariado para cada característica fue resuelto usando el software MTDFREML descrito por Boldman et al. (1995), que determina las soluciones para cada efecto fijo (b) y aleatorio (a, ap) de las ecuaciones del modelo mixto, descritas por Henderson (1984) mediante el método de máxima verosimilitud restricta libre de derivadas (Smith y Graser, 1986).
La heredabilidad y la repetibilidad así como sus respectivos errores estándar fueron obtenidos directamente usando el software MTDFREML (Boldman et al., 1995).
ESTIMACIÓN DE COMPONENTES DE (CO)VARIANZA, CORRELACIONES GENÉTICAS Y FENOTÍPICAS
Los componentes de (co)varianza y las correlaciones genéticas y fenotípicas se estimaron en la raza Holstein y en la raza Jersey, empleando un modelo animal bivariado (Mrode y Thompson, 2005). La notación matricial fue descrita como:

donde:
yl= vector de observaciones para la iésima característica (i=1 y 2, 1 corresponde a una de las características productivas (PL, PP, PG y SCS) y 2 a una de las características reproductivas (IEP, DA, NSC y TC)), obteniendo un total de 16 análisis bivariados;
bi= vector de efectos fijos para la iésima característica (los efectos fijos se definen igual como en el modelo animal univariado);
ai= vector de los efectos aleatorios genéticos aditivos para la iésima característica;
ei = vector de los efectos aleatorios del residual para la iésima característica;
Xi, Wi y Zi= matrices de incidencia que relacionan los registros de la iésima característica con los efectos fijos y aleatorios respectivamente.
Se asume que los efectos aleatorios de: ambiente permanente, genético aditivo y del residual para la iésima característica se distribuyen independientemente con media cero y varianza:

donde:
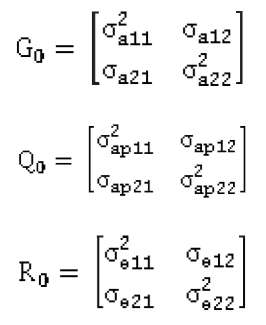

Las ecuaciones del modelo mixto para el estimador lineal insesgado (BLUE) de las funciones estimables de (b) y para el mejor predictor lineal insesgado (BLUP) de (a) y (ap) fueron definidas como:

donde:

Los análisis bivariados fueron realizados entre características con igual número de observaciones y fueron llevados a cabo mediante el uso del software MTDFREML descrito por Boldman et al. (1995), que determina las soluciones para cada efecto fijo (b) y aleatorio (a, ap) de las ecuaciones del modelo mixto, descritas por Henderson (1984) empleando el método de máxima verosimilitud restricta libre de derivadas (Smith y Graser, 1986).
Las correlaciones genéticas y fenotípicas y sus respectivos errores estándar, fueron obtenidos directamente usando el software MTDFREML (Boldman et al, 1995).
Resultados
SUMARIO ESTADÍSTICO DESCRIPTIVO
En la tabla I, se presenta el análisis descriptivo para todas las características evaluadas (PL, PG, PP, SCS, IEP, DA, NSC y TC) para Holstein y Jersey respectivamente. El promedio para PL fue mayor en la raza Holstein con 1659 litros más que el promedio para la Jersey y con valores de PG y PP de 0,8 y 0,37 % por debajo de los promedios respectivos en la raza Jersey. Para la característica SCS, el promedio fue mayor en Holstein con un valor de 4,73 comparado con el promedio obtenido en Jersey que fue de 3,98. Las características reproductivas por su parte fueron similares en las dos razas. Las características con mayor coeficiente de variación fueron: DA, NSC y PL en las dos razas, con valores de 60,5; 65,0 y 41,7 % respectivamente para la raza Holstein y 68,5; 66,0 y 39,9 % para la Jersey.
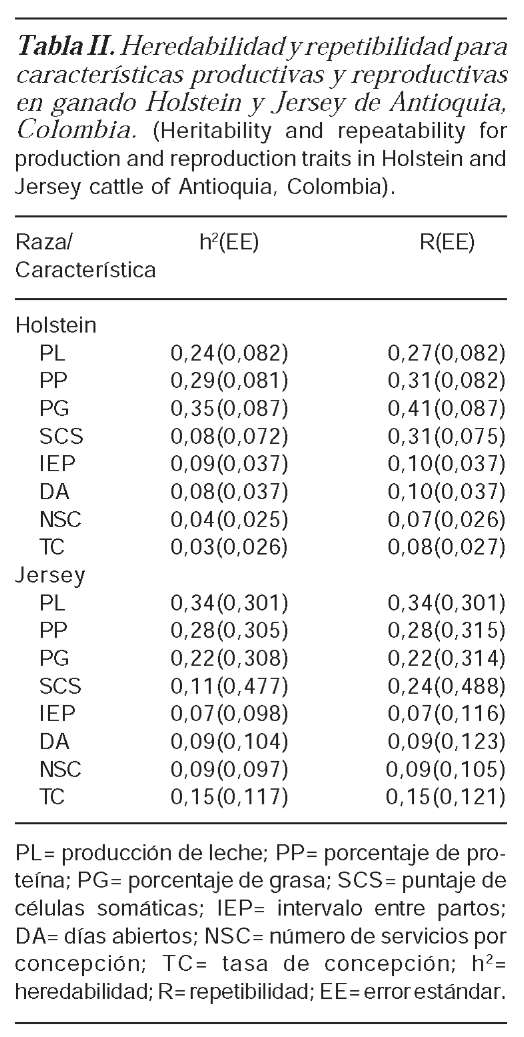
HEREDABILIDAD Y REPETIBILIDAD
Los resultados muestran que las he-redabilidades para las características reproductivas (IEP, DA, NSC y TC) y para SCS son bajas (menores de 0,15), mientras que las características productivas (PL, PP y PG) variaron en un nivel medio (entre 0,22-0,35) en Holstein y Jersey. De igual manera, la repetibilidad para características reproductivas (IEP, DA, SNC y TC) fueron bajas (menores de 0,2), mientras que las características productivas (PL, PP, PG y SCS) variaron en un nivel medio (entre 0,22-0,41) en las dos razas respectivamente. Sin embargo, en todas las características excepto en SCS, la heredabilidad y la repetibilidad fueron iguales en la raza Jersey, considerando que las varianzas obtenidas para el ambiente permanente fueron muy bajas, mientras que en la raza Holstein la repetibilidad fue ligeramente mayor que la heredabilidad. La repetibilidad solo evidenció un efecto importante del ambiente permanente en la característica SCS en las dos razas (tabla II).
CORRELACIONES GENÉTICAS Y FENOTÍPICAS
Las correlaciones genéticas más altas obtenidas para Holstein fueron: rgIEPxPL, rgIEPxPP, rgIEPxSCS y rgDAxSCS con valores de 0,77; 0,61; 0,77 y 0,73 y para Jersey: rgIEPxPP, rgIEPxSCS, rgTCxPG y rgTCxSCS con valores de 0,56; 0,49; -0,52 y -0,72 respectivamente. Los intervalos reproductivos (IEP y DA) presentaron en la mayoría de los casos asociaciones genéticas altas con todas las características productivas (PL, PP, PG y SCS), mientras que las asociaciones genéticas entre NSC y TC y las características productivas mencionadas, presentaron asociaciones genéticas bajas y moderadas en las dos razas. Por otra parte, las correlaciones fenotípicas fueron bajas en todos los casos, variaron entre -0,17 y 0,16 (tabla III).

Discusión
HEREDABILIDAD
Las heredabilidades obtenidas para características reproductivas (IEP, DA, NSC y TC) fueron bajas (menores de 0,15) para Holstein y Jersey respectivamente. En la literatura científica se reportan resultados similares en ganado lechero con hereda-bilidades que varían entre 0,01 y 0,10. M'Hamdi et al. (2010) y Ghiasi et al. (2011) reportan valores menores de 0.1 para IEP, Abe et al. (2009) y Ghiasi et al. (2011) reportan valores menores de 0,1 para DA, M'Hamdi et al. (2010) y Ghiasi et al. (2011) reportan valores muy bajos, menores de 0,05 para NSC y finalmente Abe et al. (2009) reportan valores menores de 0,06 para TC. Para SCS, al igual que en las características reproductivas mencionadas, presentó heredabilidades bajas con valores de 0,08 para Holstein y 0,11 para Jersey. VanRaden et al. (2004) y Castillo-Juárez et al. (2000) obtuvieron resultados similares (0,108 y 0,103 respectivamente). Esto confirma que las características reproductivas y de salud de la ubre son poco heredables, lo que conlleva a que el progreso genético para estas características sea bajo y que por lo tanto deben establecerse prácticas ganaderas que garanticen un buen manejo ambiental en los hatos lecheros para mejorar la eficiencia reproductiva y la salud de la ubre.
Contrario a los resultados anteriores, las características PL, PP y PG presentaron heredabilidades más altas. Para PL las here-dabilidades fueron: 0,24 para Holstein y 0,34 para Jersey. Estos valores fueron similares a los obtenidos por varios autores, que reportan resultados que oscilan entre 0,20 y 0,40 (Makgahlela et al., 2007; Abe et al., 2009; Corrales et al., 2011; Zink et al., 2012). Para PP y PG, las heredabilidades obtenidas fueron medias con valores de 0,29 y 0,35 para Holstein y 0,28 y 0,22 para Jersey. Estos valores fueron más bajos comparados con los obtenidos por Campos et al. (1994) y Makgahlela et al. (2007) quienes reportan valores mayores de 0,35. Sin embargo Abe et al. (2009) reportan heredabilidades medias para PG en la raza Holstein (0,238 en vacas de primer parto y 0,226 en vacas de segundo parto) y bajas para PP (0,186 en vacas de primer parto y 0,170 en vacas de segundo parto). Solarte y Zambrano (2012) igualmente reportaron valores bajos, menores de 0,25 para PG y PP en condiciones del trópico alto Colombiano.
CORRELACIONES GENÉTICAS
En este estudio se determinaron correlaciones genéticas entre características productivas (PL, PP, PG y SCS) y características reproductivas (IEP, DA, NSC y TC) en las razas Holstein y Jersey. Para IEP y PL, se determinaron correlaciones genéticas altas para Holstein (0,77) y bajas para Jersey (0,15). Estos resultados concuerdan con los obtenidos por varios autores que evaluaron diferentes razas lecheras. Por ejemplo Veerkamp et al. (2001); Pryce et al. (2002) y Makgahlela et al. (2007) determinaron correlaciones genéticas entre IEP y PL mayores de 0,6, mientras que Campos et al. (1994) determinaron correlaciones genéticas bajas (menores de 0,3). Un comportamiento semejante fue determinado en esta investigación para las correlaciones genéticas entre DA y PL, pero con resultados menos pronunciados, 0,40 para Holstein y 0,12 para Jersey. Resultados similares fueron encontrados por Campos et al. (1994) y Kadarmideen et al. (2003), quienes determinaron correlaciones genéticas más bajas (menores de 0,3), mientras que VanRaden et al. (2004); Abe et al. (2009) y Zink et al. (2012), obtuvieron correlaciones genéticas medias (entre 0,390,45). Sin embargo, Veerkamp et al. (2001) reportan correlaciones genéticas altas entre DA y PL (mayores de 0,6).
Debe tenerse en consideración que los DA son la parte más importante del IEP, porque la variación de la longitud de la gestación, la cual es la segunda parte del IEP es más baja. Por consiguiente las correlaciones genéticas entre IEP y PL son usualmente similares a las correlaciones genéticas entre DA y PL (Jagusiak, 2006). Estas características pueden considerarse genéticamente equivalentes, influenciadas por los mismos genes, lo que bien se conoce como pleiotropía según Falconer y Mackay, (2001). La selección para una de ellas resultará en progreso o retroceso para las demás, y en este caso la segunda opción debe ser considerada infortunadamente como uno de los factores más importantes del desequilibrio que existe entre producción y reproducción, lo que explica que vacas con altas producciones de leche presentan bajas eficiencias reproductivas. Esto sucede en la mayoría de los países que han seleccionado para PL sin tener en cuenta características de fertilidad (VanRaden et al., 2004 y Camargo, 2012).
Por otra parte, la correlación genética entre NSC y PL para la raza Holstein fue catalogado como media y de signo positivo (0,45), estos resultados son similares a los obtenidos por Veerkamp et al. (2001). De igual manera, la correlación genética entre TC y PL para la raza Holstein fue media pero de signo negativo (-0,59). Esto concuerda con los resultados obtenidos por Veerkamp et al. (2001); Abe et al. (2009) y Castillo-Juárez et al. (2000) que reportaron valores que oscilan entre -0,30 y -0,50. Estas asociaciones genéticas indeseables en ganado lechero, indican que las vacas más productivas generalmente son menos fértiles, como consecuencia del desgaste de producir altos volúmenes de leche, que desencadena una serie de problemas; principalmente una baja condición corporal y un balance energético negativo que se presenta entre la 4 y 8 semana posparto, retrasando la primera ovulación posparto y afectando el retorno al estro (Walsh et al., 2011). Sin embargo, para la raza Jersey las correlaciones genéticas (rgNSC;PL= -0,30 y rgTC;PL= 0,30) mostraron resultados favorables.
En esta investigación también fueron determinadas correlaciones genéticas entre características reproductivas (IEP, DA, NSC y TC) y características de calidad composicional de la leche (PP y PG). Para la raza Holstein las correlaciones genéticas (rgIEP;PP= 0,61; rgIEP;PG= 0,58) fueron mayores que para la raza Jersey (rgIEP;PP= 0,56; rgIEP;PG= 0,24). De manera similar, las correlaciones genéticas en la raza Holstein (rDA;PP= 0,58; r = gDA;PG = 0,39) fueron superiores que en la raza Jersey (r gDA;PP= 0,28; r gDA;PG= 0,18). Como se puede ver, en la mayoría de los casos las correlaciones genéticas obtenidas en este estudio entre características reproductivas y de calidad composicional de la leche fueron medias y de signo positivo. Campos et al. (1994) determinaron correlaciones genéticas para la raza Jersey (rgIEP;PG= 0,407; rgDA;PG= 0,253) y Toghiani (2012) para la raza Holstein (rgDA;PP= 0,741 y rgDA;PG= 0,385), que sustentan los resultados de esta investigación.
Para NSC y PP, las correlaciones genéticas obtenidas en este estudio para Holstein y Jersey fueron bajas (-0,12 y -0,27), igualmente para NSC y PG que fueron (0,07 y 0,27) respectivamente. Hermas et al. (1987) y Raheja et al. (1989) determinaron correlaciones genéticas entre NSC y PG cercanas a cero con valores que oscilan entre 0,01 y 0,11. Estos resultados sugieren que las características de calidad composicional de la leche han sido recientemente incorporadas en programas de mejoramiento genético y por tanto no presentan un fuerte antagonismo con características de fertilidad.
CORRELACIONES FENOTÍPICAS
Usando modelos bivariados, en esta investigación se obtuvieron correlaciones fenotípicas bajas entre IEP y PL con valores menores 0,1 en las dos razas (Holstein y Jersey). Estos resultados son similares a los obtenidos por Pryce et al. (2002) y Wall et al. (2003). Las correlaciones fenotípicas para DA y PL igualmente fueron bajas (menores de 0,1), resultados que fueron semejantes a los obtenidos por Yang (2009). Sin embargo, la mayoría de los autores determinaron resultados de correlaciones fenotípicas levemente superiores que oscilan entre 0,10 y 0,30 (VanRaden et al., 2004; Jagusiak, 2006 y Abe et al., 2009).
Para NSC y PL la correlación fenotípica fue 0,16 para Jersey y 0,08 para Holstein. Pryce et al. (2002); Kadarmideen et al. (2003) y Wall et al. (2003) igualmente determinaron correlaciones fenotípicas bajas para estas características (menores de 0,2) y de signo positivo, que sustentan los resultados de esta investigación. Las correlaciones fenotípicas entre TC y PL también fueron bajas pero negativas con valores de -0,17 para Holstein y -0,08 para Jersey. Estos resultados fueron similares que los obtenidos por Abe et al. (2009) y Castillo-Juárez et al. (2000) que obtuvieron valores entre -0,03 y -0,18.
Finalmente, las correlaciones fenotípicas entre características reproductivas (IEP, DA, NSC y TC) y de calidad de la leche (PP, PG y SCS) fueron bajas con valores que oscilaron entre -0,17 y 0,17. Jagusiak (2006), Yang, (2009) y Abe et al. (2009) obtuvieron resultados similares.
Conclusiones
Las heredabilidades obtenidas en este estudio confirman que las características reproductivas (IEP, DA, NSC y TC) presentan valores bajos (menores de 0,20), en las razas Holstein y Jersey y por tanto se debe considerar un buen manejo ambiental de estas características para mejorar la fertilidad de las vacas. Sin embargo las heredabilidades para características productivas (PL, PP y PG) fueron medias (0,22-0,35), excepto para SCS que fueron bajas (menores de 0,11) en las dos razas. Los resultados de heredabilidad indican que las características productivas y de calidad de la leche son mucho más heredables que las características reproductivas.
Las correlaciones genéticas entre características productivas y de calidad de la leche (PL, PP, PG y SCS) con características reproductivas (IEP, DA, NSC y TC) estuvieron dentro de una escala media y en algunos casos alta. Esto sugiere que las evaluaciones genéticas pueden realizarse mediante modelos multivariados que relacionen características productivas y reproductivas, para mejorar las precisiones de los valores genéticos de las características de baja heredabilidad como es el caso de las reproductivas.
Bibliografía
1. Abe, H.; Masuda, Y. and Susuki, M. 2009. Relationship between reproductive traits of heifers and cows and yield traits for Holsteins in Japan. J Dairy Sci, 92: 4055-4062. [ Links ]
2. Ali, A.K.A. and Shook, G.E. 1980. An optimun transformation for somatic cell concentration in milk. J Dairy Sci, 63: 487-490. [ Links ]
3. Boldman, K.G.; Kriese, L.A.; Van Vleck, L.D.; Van Tassell, C.P. and Kachman, S.D. 1995. A manual for use of MTD-FREML: A set of programs to obtain estimates of variances and covariances. USDA. ARS. Washington, DC. [ Links ]
4. Camargo, O. 2012. La vaca lechera: Entre la eficiencia económica y la ineficiencia biológica. Arch Zootec, 61: 13-29. [ Links ]
5. Campos, M.S.; Wilcox, C.J.; Becerril, C.M. and Diz, A. 1994. Genetic parameters for yield and reproductive traits of Holstein and Jersey cattle in Florida. J Dairy Sci, 77: 867-873. [ Links ]
6. Castillo-Juárez, H.; Oltenacu, P.A.; Blake, R.W.; Mcculloch, C.E. and Cienfuegos-Rivas, E.G. 2000. Effect of herd environment on the genetic and phenotypic relationships among milk yield, conception rate and SCS in Holstein cattle. J Dairy Sci, 83: 807-814. [ Links ]
7. Corrales, J.; Cerón-Muñoz, M.; Cañas, J.; Herrera, C. y Calvo, S. 2011. Parámetros genéticos de características de tipo y producción en ganado Holstein del Departamento de Antioquia. Rev MVZ. Córdoba, 17: 2870-2877. [ Links ]
8. Echeverri, J.; López, A. and Parra, J.E. 2010. Software control 1 Manejo y control de producción para hatos lecheros. Laboratorio de Mejoramiento Genético Animal. Universidad Nacional de Colombia. [ Links ]
9. Falconer, D.S. y Mackay, T.F. 2001. Introducción a la genética cuantitativa. 4a. ed. Editorial Acribia. Zaragoza. España. [ Links ]
10. Ghiasi, H.; Pakdel, A.; Nejati-Javaremi, A.; Mehrabani-Yeganeh, H.; Honarvar, M.; González-Recio, O.; Carabaño, M.J. and Alenda, R. 2011. Genetic variance components for female fertility in Iranian Holstein cows. Livest Sci, 139: 277-280. [ Links ]
11. Henderson, C.R. 1984. Applications of linear models in animal breeding. Canadian Catalog Publication Data. Univ Guelph. Canada, USA. [ Links ]
12. Henderson, C.R. and Quaas, R.L. 1976. Multiple trait evaluation using relatives' records. J Anim Sci, 43: 1188-1197. [ Links ]
13. Hermas, S.A.; Young, C.W. and Rust, J.W. 1987. Genetic relationships and additive genetic variation of productive and reproductive traits in Guernsey dairy cattle. J Dairy Sci, 70: 1252-1257. [ Links ]
14. Jagusiak, W. 2006. Fertility measure in Polish Black-and-White cattle. Phenotypic and genetic correlations between fertility measures and milk productions traits. J Anim Feed Sci, 15: 371-380. [ Links ]
15. Jamrozik, J.; Fatehi, J.; Kistemaker, G.J. and Schaeffer, L.R. 2005. Estimates of genetic parameters for Canadian Holstein female reproduction traits. J Dairy Sci, 88: 2199-2208. [ Links ]
16. Kadarmideen, H.N.; Thompson, R.; Coffey, M.P. and Kossaibati, M.A. 2003. Genetic parameters and evaluations from single - and multiple-trait analysis of dairy cow fertility and milk production. Livest Prod Sci, 81: 183-195. [ Links ]
17. M'Hamdi, N.; Aloulou, R.; Brar, S.K.; Bouallegue, M. and Ben Hamouda, M. 2010. Phenotypic and genetic parameters of reproductive traits in Tunisian Holstein Cows. Biotech Anim Husb, 26: 297-307. [ Links ]
18. Makgahlela, M.L.; Banga, C.B.; Norris, D.; Dzama, K. and N'gambi, J.W. 2007. Genetic correlations between female fertility and production tratis in south African Holstein Cattle. S Afr J Anim Sci, 37: 180-187. [ Links ]
19. Miglior, F.; Muir, B.L. and Van Doormaal, B.J. 2005. Selection indices in Holstein cattle of various countries. J Dairy Sci, 88: 1255-1263. [ Links ]
20. Mrode, R. and Thompson R. 2005. Linear models for the prediction of animal breeding values. 2nd ed. CAB Publishing. Oxford, UK. [ Links ]
21. Pryce, J.E.; Coffey, M.P.; Brotherstone, S.H. and Woolliams, J.A. 2002. Genetic relationships between calving interval and body conditions score conditional on milk yield. J Dairy Sci, 85: 1590-1595. [ Links ]
22. Raheja, K.L.; Burnside, E.B. and Schaeffer, L.R. 1989. Relationships between fertility and production in Holstein dairy cattle in different lactations. J Dairy Sci, 72: 2670-2678. [ Links ]
23. SAS®. 2006. SAS/STAT. User's Guide. Statistical Analysis Systems Institute. Version 9.1th ed. SAS Institute Inc. Cary, NC. [ Links ]
24. Sbardella, M. and Gaya, L.G. 2010. Unfavourable side implications of animal breeding in livestock species. Arch Zootec, 59: 157-168. [ Links ]
25. Sewalem, A.; Kistemaker, G.J. and Miglior, F. 2010. Relationship between female fertility and production traits in Canadian Holstein. J Dairy Sci, 93: 4427-4434. [ Links ]
26. Smith, S.P. and Graser, H.U. 1986. Estimating variance components in a class of mixed models by restricted máximum likelihood. J Dairy Sci, 69: 1156-1165. [ Links ]
27. Solarte, C.E. and Zambrano, G.L. 2012. Characterization and genetic evaluation of Holstein cattle in Nariño, Colombia. Rev Col Cienc Pec, 25: 539-547. [ Links ]
28. Toghiani, S. 2012. Genetic relationships between production traits and reproductive performance in Holstein dairy cows. Arch Tierz, 55: 458-468. [ Links ]
29. Van Raden, P.M.; Sanders, A.H.; Tooker, M.E.; Miller, R.H.; Norman, H.D.; Kuhn, M.T. and Wiggans, G.R. 2004. Development of a national genetic evaluation for cow fertility. J Dairy Sci, 87: 2285-2292. [ Links ]
30. Veerkamp, R.F.; Koenen, E.P.C. and De Jong, G. 2001. Genetic correlations among body condition score, yield, and fertility in first-parity cows estimated by random regression models. J Dairy Sci, 84: 2327-2335. [ Links ]
31. Wall, E.; Brotherstone, S.; Woolliams, J.A.; Banos, G. and Coffey, M.P. 2003. Genetic evaluation of fertility using direct and correlated traits. J Dairy Sci, 86: 4093-4102. [ Links ]
32. Walsh, S.W.; Williams, E.J. and Evans, A.C.O. 2011. A review of the causes of poor fertility in high milk producing dairy cows. Anim Reprod Sci, 123: 127-138. [ Links ]
33. Yang, L. 2009. Phenotypic relatioships between milk protein percentage, reproductive performance and body condition score in Irish dairy cattle. Thesis MSc. Massey University. Palmerston North. New Zelanda. [ Links ]
34. Zink, V.; Lassen, J. and tfpková, M. 2012. Genetic parameters for female fertility and milk productions traits in first-parity Czech Holstein cows. Czech J Anim Sci, 57: 108-114. [ Links ]
Recibido: 23-8-13.
Aceptado: 20-3-14

