










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkNutrición Hospitalaria
versión On-line ISSN 1699-5198versión impresa ISSN 0212-1611
Nutr. Hosp. vol.28 no.6 Madrid nov./dic. 2013
https://dx.doi.org/10.3305/nh.2013.28.6.6790
ORIGINAL / Investigación animal
Effects of human milk on blood and bone marrow cells in a malnourished mice model; comparative study with cow milk
Efecto de la leche humana sobre células de sangre y de médula ósea en un modelo de ratos desnutridos; estudio comparativo con leche de vaca
Isabel García1, Susana Salva2, Hortensia Zelaya1, Julio Villena2 and Graciela Agüero1
1Instituto de Bioquímica Aplicada. Facultad de Bioquímica, Química y Farmacia. Universidad Nacional de Tucumán. Argentina.
2Centro de Referencia para Lactobacilos (CERELA-CONICET). Argentina.
This work was supported by grants from CIUNT 26 D/303.
ABSTRACT
Introduction: It has been demonstrated that the alterations caused by nutrient deficiency can be reverted by adequate nutritional repletion.
Objective: To perform comparative studies between human and cow milks in order to evaluate the impact of both milks on the recovery of blood and bone marrow cells affected in malnourished mice.
Method: Weaned mice were malnourished after consuming a protein free diet for 21 days. Malnourished mice received cow or human milk (CM or HM) for 7 or 14 consecutive days. During the period of administration of milk, the mice consumed the protein free diet ad libitum. The malnourished control (MNC) group received only protein free diet whereas the well-nourished control (WNC) mice consumed the balanced conventional diet.
Results and Discussion: Both milks normalized serum albumin levels and improved thymus weight. Human milk was less effective than cow milk to increase body weight and serum transferrin levels. In contrast, human milk was more effective than cow milk to increase the number of leukocytes (WNC: 6.90 ± 1.60a; MNC: 2.80 ± 0.90b; CM 7d: 3.74 ± 1.10b; HM 7d: 7.16 ± 1.90a; CM 14d: 4.35 ± 1.20b; HM 14d: 6.75 ± 1.20a (109/L);p < 0.05) and lymphocytes (WNC: 5.80 ± 0.36a; MNC: 1.80 ± 0.40b; CM 7d: 2.50 ± 0.30b; HM 7d: 4.20 ± 0.50c; CM 14d: 3.30 ± 0.31d; HM 14d: 4.70 ± 0.28c (109/L);p < 0.05) in peripheral blood. Both milks induced an increment in mitotic pool cells in bone marrow and α-naphthyl butyrate esterase positive cells in peripheral blood. They also normalized phagocytic function in blood neutrophils and oxidative burst in peritoneal cells.
Conclusion: Both milks were equally effective to exert favorable effects on the number of the bone marrow cells and the functions of the blood and peritoneal cells involved in immune response. However, only human milk normalized the number of leukocytes and increased the number of neutrophils in peripheral blood.
Key words: Malnourished mice. Human milk. Cow milk. Blood cells. Bone marrow cells.
RESUMEN
Introducción: Las alteraciones causadas por la deficiencia de nutrientes pueden ser revertidas por un aporte nutricional adecuado.
Objetivos: Realizar estudios comparativos entre leche humana y leche de vaca para evaluar su impacto en la recuperación de las células de sangre y de médula ósea afectadas en ratones desnutridos.
Métodos: Los ratones fueron desnutridos al recibir una dieta libre de proteínas durante 21 días a partir del destete. Posteriormente, estos ratones desnutridos recibieron leche de vaca (LV) o leche humana (LH) durante 7 o 14 días consecutivos, mientras continuaban consumiendo la dieta libre de proteínas ad libitum. El grupo control de desnutrición (CD) sólo recibió la dieta libre de proteínas mientras que los ratones controles bien nutridos (CBN) consumieron la dieta balanceada convencional.
Resultados y Discusión: Ambas leches normalizaron los niveles de albumina sérica e incrementaron el peso del timo. La leche humana fue menos efectiva que la leche de vaca para incrementar el peso corporal y los niveles de transferrina en suero. Sin embargo, la leche humana fue más efectiva para incrementar el número de leucocitos (CBN: 6,90 ± 1,60a; CD: 2,80 ± 0,90b; LV 7d: 3,74 ± 1,10b; LH 7d: 7,16 ± 1,90a; LV 14d: 4,35 ± 1,20b; LH 14d: 6,75 ± 1,20a (109/L);p < 0,05) y linfocitos (CBN: 5,80 ± 0,36a; CD: 1,80 ± 0,40b; LV 7d: 2,50 ± 0,30b; LH 7d: 4,20 ± 0,50c; LV 14d: 3,30 ± 0,31d; LH 14d: 4,70 ± 0,28c (109/L); p < 0,05) en sangre periférica. Ambas leches indujeron un incremento de las células del compartimiento mitótico de médula ósea y de las células ±-naftil butirato esterasa positivas en sangre periférica. Además, normalizaron la función fagocítica en neutrófilos de sangre periférica y el estallido oxidativo en las células peritoneales.
Conclusiones: Ambas leches fueron igualmente efectivas para ejercer efectos favorables en el número de las células de la médula ósea y en las funciones de las células peritoneales y de la sangre involucradas en la respuesta inmune. Sin embargo, sólo la leche humana normalizó el número de leucocitos e incrementó el número de neutrófilos en sangre periférica.
Palabras clave: Ratones desnutridos. Leche humana. Leche de vaca. Células sanguíneas. Célula de la médula ósea.
Abbreviations
αNBE+: α-naphthyl butyrate esterase positive cells.
βG+: β-glucuronidase positive cells.
BCD: balanced conventional diet.
CM: cow milk.
CM 7d: malnourished mice replete with CM for 7 consecutive days.
CM 14d: malnourished mice replete with CM for 14 consecutive days.
Hb: haemoglobin concentration.
HCT: haematocrit.
HM: human milk.
HM 7d: malnourished mice replete with HM for 7 consecutive days.
HM 14d: malnourished mice replete with HM for 14 consecutive days.
MNC: malnourished control mice.
MPO: myeloperoxidase.
NBT: nitro blue tetrazolium.
NBT+: NBT positive cells.
PBS: phosphate buffer saline.
PFD: protein free diet.
WNC: well-nourished control mice.
Introduction
Protein energy deprivation alters cellular immunity, phagocyte function, the complement system, secretory immunoglobulin A concentrations, and cytokine production1. As a consequence, malnutrition increases the susceptibility to infections. It was reported that protein energy deprivation also causes severe lesions in organs with high cellular proliferation such as the intestinal epithelium2 and the hematopoietic tissue3.
It has been demonstrated that the alterations caused by nutrient deficiency can be reverted by adequate nutritional repletion. Numerous experiences suggest that milk is superior to other repletion diets used in the treatment of malnutrition in terms of mortality, sepsis, improvement of intestinal permeability and weight gain4.
Human milk is a bodily fluid which, apart from being an excellent nutritional source for the growing infant, also contains a variety of immune components such as antibodies, growth factors, cytokines, antimicrobial compounds, and specific immune cells5-7. These factors help to support the immature immune system of newborns and to protect them against infectious risks during the postnatal period while its own immune system matures8. More recent clinical and experimental observations also suggest that human milk not only provides passive protection, but also can directly modulate the immunological development of the recipient infant9. In addition, the presence of growth factors and hormones, both beneficial for the host, has been observed in milk from different species, including human and bovine10-12. Proteins in human milk not only provide amino acids but also bind to and facilitate the absorption of nutrients, stimulate the growth and development of the intestinal epithelium, and aid in the digestion of other nutrients13.
On the other hand, cow milk is one of the most important sources of dietary protein in humans. It is constituted by high protein quality arises both from its nutritional value and its physiological properties. Physiological properties of cow milk protein include acute regulatory effects on nutrient bioavailability or on immune mechanisms and longer term potential benefits for cardiovascular-system or tissue development14.
Considering these relevant antecedents, in the present work it was performed comparative studies between human and cow milks in order to evaluate the impact of both milks on the recovery of blood and bone marrow cells affected in malnourished mice.
Materials and methods
Animals
Balb/c mice were obtained from the closed colony kept at the bioterium of CERELA. They were housed in plastic cages in a controlled atmosphere (temperature 22 ± 2o C; humidity 55 ± 2%) with a 12 h light/dark cycle.
Feeding procedures
Weaned mice (3-week-old) were malnourished after they consumed a protein-free diet (PFD) for 21 days.
Milk from creole cows (CM) was provided by farmers from Tucumán, Argentina. Human milk (HM) was obtained from healthy women with normal pregnancy and delivery. Informed consent was obtained from every mother. Exclusion criteria included gestation < 37 weeks, birth weight < 2.5 kg, multiple pregnancy, major illness requiring intensive care admission, and major congenital anomalies. The samples of mature milks were expressed manually from each breast, collected in sterile containers between 9.00-11.30 a.m. and then aliquoted. Samples were stored at -70o C until use.
Malnourished mice received CM or HM for 7 (CM 7d, HM 7d) or 14 (CM 14d, HM 14d) consecutive days. During the period of administration of milk, the mice consumed the PFD ad libitum. The malnourished control (MNC) group received only PFD whereas the well-nourished control (WNC) mice consumed the balanced conventional diet (BCD).
Samples from control (WNC and MNC), CM 7d, HM 7d, CM 14d, and HM 14d groups were obtained at the end of each feeding period.
Animals were cared for in accordance with standard guidelines (Canadian Council on Animal Care, 1993). The experimental protocol was approved by the Ethical Committee for Animal Care of CERELA and of the Universidad Nacional de Tucumán, Argentina.
Study of nutritional state
Body weight and thymus weight were determined at the beginning and end of each feeding period with an electronic balance with a sensitivity of 0.01 g and 0.001 g respectively. Body and thymus weight resulted from the mean of the values obtained in three different weighings performed alternately. Body weight was expressed as percentage of body weight increase with respect to the initial value.
Thymus weight was expressed in mg and could only be determined in the animals that received the milk supplementation since malnutrition caused extreme involution of the organ.
For the determination of serum parameters, blood samples were obtained by cardiac puncture from sodium pentobarbital-anesthetized animals. The samples were collected in glass tubes and the serum was obtained.
Albumin concentration was determined using a colorimetric assay based on albumin binding to bromo-cresol green (Wiener Lab, Rosario, Argentina).
Serum transferrin was determined by radial immunodiffusion (Diffu-Plate; Biocientífica, S.A., Buenos Aires, Argentina).
Study of blood cells
Basic haematological parameters: blood samples were collected in tubes containing EDTA as an anticoagulant. Haemoglobin concentration (Hb) was carried out using the haemiglobincyanide method. Haematocrit (HCT) was determined manually by micro-haematocrit techniques. Total number of blood leukocytes was determined with a haemocytometer. The results were expressed as 109/L. Differential blood leukocyte count was determined by counting 200 cells in blood smears stained with May Grünwald-Giemsa using a light microscope (100x) and absolute numbers were calculated.
Study of bone marrow cells
Anesthetized mice were killed by cervical dislocation and bone marrow samples were obtained by flushing the femoral cavity with phosphate buffer saline (PBS).
Differential cell counts were carried out by counting 500 cells in bone marrow smears stained with May Grunwald-Giemsa. Myeloid cells were grouped into the mitotic pool, which includes cells capable of replication (myeloblasts, promyelocytes and myelocytes), and the post-mitotic pool, whose cells usually do no replicate but are able to evolve towards more mature and differentiated cells (metamyelocytes, band cells and neutrophils). Lymphoid cells and erythroblasts were also counted. The results of myeloid cells, lymphoid cells, and erythroblast counts were expressed as percentages of total white bone marrow cells.
To study T cells maturation, cytochemical assays were performed according the maturation scheme proposed by Basso et al.15 It was determined the percentage of β-glucuronidase positive (βG+) cells and α-naphthyl butyrate esterase positive (αNBE+) cells in bone marrow and blood samples using commercial cytochemical assay kits (Sigma-Aldrich, St Louis, MO, USA). The cells were counted under a light microscope (100x) and were regarded as positive cells that showed granular localized coarse positivity.
Study of myeloperoxidase (MPO) activity in blood neutrophils and bone marrow myeloid cells
Myeloperoxidase (MPO) activity was determined using a cytochemical method (Washburn test) with benzidine as a myeloperoxidase chromogen. Cells were graded as negative or weakly, moderately or strongly positive and were used to calculate the score.
Study of nitro blue tetrazolium (NBT) reduction test in peritoneal macrophages
The oxidative burst was studied in peritoneal lavage-derived macrophages using the nitro blue tetrazolium (NBT) reduction test (Sigma-Aldrich, St Louis, MO, USA). Samples were examined under a light microscope for blue precipitate. A hundred cells were counted and the percentage of NBT positive cells (NBT+) was determined.
Statistical analysis
Experiments were performed in triplicate. For each experiment, 30-36 mice were used (5-6 animals each group). Results were expressed as means ± SD. After verification of the normal distribution of data, ANOVA was used. Tukeyis test (for pairwise comparisons of the means) was used to test for differences between the groups. Differences were considered significant at p < 0.05.
Results
Nutritional parameters
The body weight of mice in the MNC group was significantly lower than in the WNC group (Table I). Only the feeding with CM induced a significant increase in the body weight with respect to initial values (Table I).
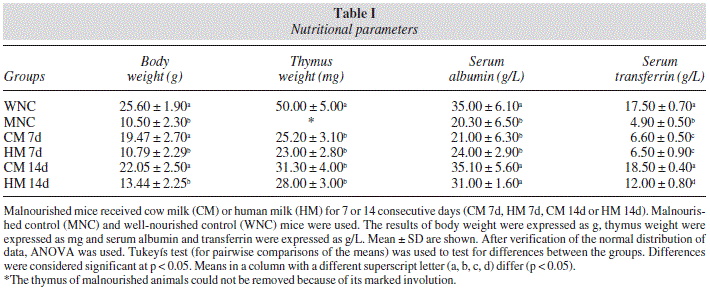
The thymus of malnourished animals could not be removed because of its marked involution. Feeding with HM or CM for 7 and 14 d were equally effective to increase thymus weight, although the values of the well-nourished control were not reached (Table I).
Malnutrition induced a significant decrease in serum albumin levels (p < 0.05). Both milks administered for 14 d were effective to normalize this parameter (Table I). In addition, protein deficiency induced a significant decrease in serum transferrin levels (p < 0.05). HM and CM administration for 14 d induced a significant recovery of serum transferrin levels, although only the animals that received CM reached normal values (Table I).
Study of blood cells
Protein deficiency decreased Hb concentration and HCT (p < 0.05). The administration of both milks for 7 d was sufficient to normalize Hb and HCT values (Figs. 1A and B).

Malnutrition induced a significant decrease in number of total leukocytes and lymphocytes in peripheral blood (Table II). Administration of HM for 7 d normalized total leukocyte numbers, while CM group reached normal values at 14 d (Table II).
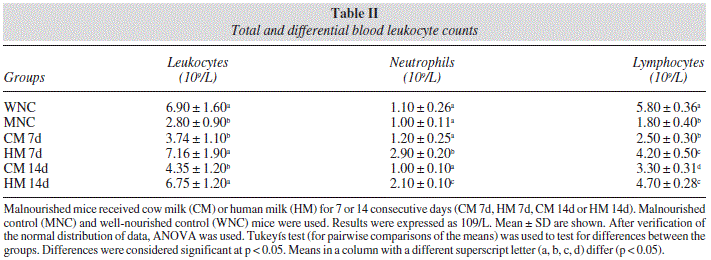
The number of lymphocytes increased after repletion with both milks, however HM was more efficient than CM to recover this cell population (Table II). In addition, neutrophil numbers were increased only after HM administration.
Study of bone marrow cells
The MNC group showed a significant decrease (p < 0.05) in erythroblast counts. Both milks administered for 14 d increased the percentage of these cells above the values of the WNC (Fig. 1C). In addition, protein deprivation induced a significant decrease in mitotic pool cells, which were normalized by both milks (Fig. 2A). The post-mitotic pool was not significantly modified by malnourishment (Fig. 2B). The percentage of lymphoid cells in bone marrow was also decreased in the MNC group. Both milks administered for 14 d normalized the number of these cells (Fig. 2C).

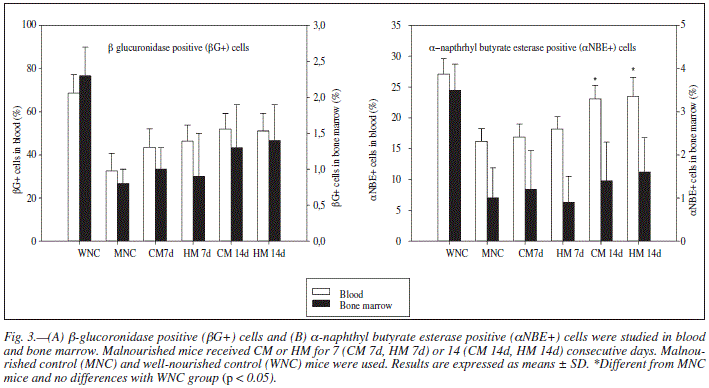
The MNC group showed a significant decrease in the percentage of βG+ cells and αNBE+ cells in blood and bone marrow. Both milks administered for 14 d induced a significant recovery in the number of αNBE+ cells in peripheral blood (Figs. 3A and B).
Study of MPO activity in blood neutrophils and bone marrow myeloid cells
The MNC group showed a decrease (p < 0.05) in MPO positive blood neutrophils. Both milks induced a statistically significant improvement when administered for 14 d, values similar to those of the WNC being reached (Fig. 4A). Malnourished animals showed a high percentage of neutrophil leukocytes with weak MPO positivity. After 14 d of feeding, a high percentage of neutrophil leukocytes with strong MPO positivity were observed (Fig. 4B).

Malnutrition did not induce significant alterations in MPO activity in bone marrow (Fig. 4A).
Study of NBT reduction test in peritoneal macrophages
The respiratory burst, measured with the NBT test, was significantly decreased by malnutrition (p < 0.05). Feeding with the different milks for 7 or 14 d caused increases of NBT+ cells above the values of the WNC (Fig. 4C).
Discussion
Human milk is considered the ideal food for the newborn during the first months of life16. In addition, it is amply documented that milk is an important source of proteins with high nutritional value17,18. To evaluate the nutritional status in this experimental model it was determined body weight, albumin and serum transferrin. It is known that a large number of cytokines present in human milk exert their beneficial biological activities mainly at the level of the digestive tract19. Thus, the recovery of the gastrointestinal tract, which is severely damaged in malnourished hosts20, would favour the adequate absorption of nutrients, enabling the effect of bioactive metabolites to be exerted at the level of other organs. However, in this experimental model, HM did not lead to significant increase in body weight, which could be attributed to its lower protein content compared with CM. Weight gain is mainly associated with protein intake as was reported by Berkey et al21. and Barr et al18. Malnutrition also caused a significant decrease in albumin and serum transferrin. Similar results were found by Rahman et al22. in malnourished children. Nutritional repletion with HM or CM for 14 d were equally efficient to normalize serum albumin values. Nevertheless, HM did not normalize transferrin values, while CM induced an increase of this serum protein, reaching values similar to those in the WNC group. The different behaviour of HM and CM on serum albumin and transferrin values could be attributed to the greater nutritional capacity of CM as a consequence of its greater protein content23.
Malnutrition induced a significant decrease in numbers of leukocytes and lymphocytes in peripheral blood, an effect that could be attributed to the atrophy of organs such as thymus and bone marrow24.
When it was studied the effect of the protein deficient diet on thymus, it was observed a marked involution. Thymic atrophy in malnourished hosts has been attributed to decreased levels of leptin and high concentrations of glucocorticoids in serum25. Glucocorticoids probably increase apoptosis, as demonstrated by Hughes et al. in in vitro assays with rat lymphocytes26. On the other hand, there is in vitro evidence suggesting that leptin can activate cytokine signalling pathways in T cells, thereby regulating their proliferation and apoptosis27. Feeding with HM or CM for 7 and 14 d were equally effective to increase thymus weight, although the values of the WNC were not reached.
It has been shown that malnutrition affects tissues that have a high turnover rate and cell proliferation as the hematopoietic tissue, inducing an impairment of blood cell production, leading to hypoplasia and structural changes of bone marrow28,29. In this work, it was observed that malnutrition induced a decrease in the number of total bone marrow cells, including granulocytes, erythroblasts, and lymphoid cells. These results agree with previous studies that showed bone marrow atrophy with reduced number of leukocytes, granulocytes and lymphocytes in malnourished mice29,30.
Peripheral manifestation of the activity of bone marrow after milk repletion allowed to conclude that HM was more effective than CM to recover normal values of leukocytes and to increase the number of lymphocytes. This phenomenon would probably be mediated by growth factors and/or cytokines contained in human milk31. In this sense, Bryan et al. reported that human milk provides IL-2, which participates in lymphoid differentiation and in the regulation of T lymphocytes32.In addition, it was demonstrated that milk proteins such as β-casein are able to stimulate cell proliferation in cultures of human immunocompetent cell lines32.
The decrease in βG+ and αNBE+ cells in peripheral blood and bone marrow in malnourished animals would suggest deficiencies in T cell maturation. Both milks increased the numbers of αNBE+ cells in peripheral blood, and therefore, they would improve the maturation stage of lymphocytes.
HM administered for 7 or 14 d also caused an increase in neutrophils in peripheral blood and in pool mitotic cells in bone marrow. This phenomenon could be interpreted on the basis of the works of Sinha et al33., who reported that HM contains hematopoietic stimulatory activity that would be protected from gastric digestion by anti-proteolytic agents present in the milk.
Multiple abnormalities of immune response were described as consequence of decreased phagocytosis induced by malnutrition34. In the present work, it was observed that both milks allowed the recovery of the phagocytic and bactericidal activity of peritoneal macrophages in malnourished mice after 7 d of administration. These results are supported by the findings of other investigators, who reported that a lactoferrin-derived peptide increased the phagocytic activity of human neutrophils, binding directly to them or exerting opsonin activity35. Moreover, Li et al. reported that a glycomacropeptide derived from bovine milk increased the phagocytic activity of a human cell line36. Therefore, several components of milks would be responsible for the recovery of phagocytes activities in malnourished mice.
Conclusion
In conclusion, both milks were equally effective to exert favourable effects on the number of the bone marrow cells and the functions of the blood and peritoneal cells involved in immune response. However, only HM normalized the number of total leukocytes and increased the number of neutrophils in peripheral blood. This would be the consequence of thymus and bone marrow improvement.
Acknowledgements
The authors also wish to thank Wiener Lab, Rosario, Argentina for their kind donation of assay kits.
Author disclosure statement
The authors declare that they have no competing financial interests.
References
1. Scrimshaw NS, SanGiovanni JP. Synergism of nutrition, infection, and immunity: an overview. Am J Clin Nutr 1997; 66: 464-77. [ Links ]
2. Deo M, Meera M, Ramalingaswami V. Cell generation in protein deficiency. Nature 1967; 216: 499. [ Links ]
3. Borelli P, Mariano M, Borojevic R. Protein malnutrition: Effect on myeloid cell production and mobilization into inflammatory reactions in mice. Nutrition Research 1995; 15: 1477-85. [ Links ]
4. Scherbaum V, Furst P. New concepts on nutritional management of severe malnutrition: the role of protein. Curr Opin Clin Nutr Metab Care 2000; 3: 31-8. [ Links ]
5. Goldman AS. The immune system in human milk and the developing infant. Breastfeed Med 2007; 2: 195-204. [ Links ]
6. Hosea Blewett HJ, Cicalo MC, Holland CD, Field CJ. The immunological components of human milk. Adv Food Nutr Res 2008; 54: 45-80. [ Links ]
7. Lawrence RM, Pane CA. Human breast milk: current concepts of immunology and infectious diseases. Curr Probl Pediatr Adolesc Health Care 2007; 37: 7-36. [ Links ]
8. Paramasivam K, Michie C, Opara E, Jewell AP. Human breast milk immunology: a review. Int J Fertil Womens Med 2006; 51: 208-17. [ Links ]
9. Garofalo R. Cytokines in human milk. J Pediatr 2010; 156: 36-40. [ Links ]
10. Koldovsky O. Hormones in milk: their possible physiological significance for the neonate. In: Lebenthal L, (ed.). In: Gastroenterology and nutrition in infancy Anonymous New York: Raven Press 2002. pp. 97-119. [ Links ]
11. Meisel H, Bockelmann W. Bioactive peptides encrypted en mil proteins: proteolytic activation and thropho-functional properties. Antonie Van Leeuwenhoek 1999; 76: 207-15. [ Links ]
12. Matar C, Goulet J, Bernier R, Brochu E. Bioactive peptides from fermented foods: their role in the immune system In: Fuller R and Perdigón G, (eds.). Probiotics 3: Immunomodulation by the gut microflora and probiotics Anonymou London: Kluwer Academis Publisher, 2000. pp. 193-212. [ Links ]
13. Lonnerdal B. Biochemistry and physiological function of human milk proteins. Am J Clin Nutr 1985; 42: 1299-317. [ Links ]
14. Bos C, Gaudichon C, Tome D. Nutritional and physiological criteria in the assessment of milk protein quality for humans. J Am Coll Nutr 2000; 19: 191S-205S. [ Links ]
15. Basso G, Cocito M, Semenzato G, Pezzuto A, Zanesco L. Cytochemical study of thymocytes and T lymphocytes. Br J Haematol 1980; 4: 577-82. [ Links ]
16. Committee on Nutritional Status During P, Lactation and Institute of M. Nutrition During Lactation. The National Academies Press, 1991. [ Links ]
17. Donnen P, Brasseur D, Dramaix M, Assimbo V, Hennart P. Effects of cow's milk supplementation on milk output of protein deficient lactating mothers and on their infants' energy and protein status. Trop Med Int Health 1997; 1: 38-46. [ Links ]
18. Barr SI, McCarron DA, Heaney RP, et al. Effects of increased consumption of fluid milk on energy and nutrient intake, body weight, and cardiovascular risk factors in healthy older adults. J Am Diet Assoc 2002; 102: 1672-4. [ Links ]
19. Field C. The immunological components of human milk and their effect on immune development in infants. J Nutr 2005; 135: 1-4. [ Links ]
20. Allori C, Agüero G, de Ruiz Holgado AP, de Nader OM, Perdigón G. Gut mucosa morphology and microflora changes in malnourished mice after renutrition with milk and administration of Lactobacillus casei. J Food Prot 2000; 63: 83-90. [ Links ]
21. Berkey CS, Rockett HR, Willett WC, Colditz GA. Milk, dairy fat, dietary calcium, and weight gain: a longitudinal study of adolescents. Arch Pediatr Adolesc Med 2000; 159: 543-50. [ Links ]
22. Rahman MZ, Begum BA. Serum total protein, albumin and A/G ratio in different grades of protein energy malnutrition. Mymensingh Med J 2005; 14: 38-40. [ Links ]
23. Winkler MF, Pomp A, Caldwell MD, Albina JE. Transitional feeding: the relationship between nutritional intakeand plasma protein concentrations. J Am Diet Assoc 1989; 89: 969-70. [ Links ]
24. Savino W. The thymus gland is a target in malnutrition. Eur J Clin Nutr 2002; 56 Supl. 3: 46-9. [ Links ]
25. Ahima RS, Prabakaran D, Mantzoros C, et al. Role of leptin in the neuroendocrine response to fasting. Nature 1996; 382: 250-2. [ Links ]
26. Hughes F, Cidlowski J. Glucocorticoid-induced thymocyte apoptosis: Protease-dependent activation of cell shrinkage and DNA degradation. J Steroid Biochem Mol Biol 1998; 65: 207-17. [ Links ]
27. Palmer G, Aurrand-Lions M, Contassot E, et al. Indirect effects of leptin receptor deficiency on lymphocyte populations and immune response in db/db mice. J Immunol 2006; 177: 2899-907. [ Links ]
28. Salva S, Villena J, Racedo S, Alvarez S, Agüero G. Lactobacillus casei addition to a repletion diet-induced early normalisation of cytokine profils during a pneumococcal infection in malnourished mice. Food and Agricultural Immunology 2008; 19:195-211. [ Links ]
29. Vinolo MA, Crisma AR, Nakajima K, Rogero MM, Fock RA, Borelli P. Malnourished mice display an impaired hematologic response to granulocyte colony-stimulating factor administration. Nutr Res 2008; 28: 791-7. [ Links ]
30. Fock RA, Blatt SL, Beutler B, et al. Study of lymphocyte subpopulations in bone marrow in a model of protein-energy malnutrition. Nutrition 2009; 26: 1021-8. [ Links ]
31. Castellote C, Casillas R, Ramirez-Santana C, et al. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J Nutr 2011; 141: 1181-7. [ Links ]
32. Bryan D, Forsyth K, Gibson R, Hawkes J. Interleukin-2 in human milk: a potential modulator of lymphocyte development in the breastfed infant. Cytokine 2006; 33: 289-93. [ Links ]
33. Sinha S, Yunis A. Isolation of colony stimulating factor from milk. Biochem Biophys Res Commun 1983; 114: 797-803. [ Links ]
34. Villena J, Racedo S, Agüero G, Bru E, Medina M, Alvarez S. Lactobacillus casei improves resistance to pneumococcal respiratory infection in malnourished mice. J Nutr 2005; 135: 1462-9. [ Links ]
35. Miyauchi H, Hashimoto M, Nakajima I, Shinoda Y, Fukuwatari Y, Hayasawa H. Bovine lactoferrin stimulates the phagocytic activity of human neutrophils: identification of its active domain. Cell Immunol 1998; 187: 34-7. [ Links ]
36. Li EW, Mine Y. Immunoenhancing effects of bovine glycomacropeptide and its derivatives on the proliferative response and phagocytic activities of human macrophagelike cells, U937. J Agric Food Chem2004; 52: 2704-8. [ Links ]
![]() Correspondence:
Correspondence:
Graciela Agüero.
Instituto de Bioquímica Aplicada.
Facultad de Bioquímica, Química y Farmacia.
Universidad Nacional de Tucumán.
Balcarce, 747.
4000 San Miguel de Tucumán. Argentina.
E-mail: gaguero@fbqf.unt.edu.ar
Recibido: 17-IV-2013.
1.a Revisión: 24-VI-2013.
Aceptado: 18-IX-2013.

