










 Servicios personalizados
Servicios personalizados
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
El metabolismo basal está regulado por el hipotálamo a través de un complejo circuito neuronal que controla la cantidad de energía que se ingiere y utiliza en un momento dado. Los componentes importantes de este sistema incluyen la sensación de hambre y de saciedad, la actividad del sistema nervioso autónomo y el sistema endocrino (1). La dieta juega un papel clave en la composición corporal pero, además de afectar al balance energético, todavía existe información limitada sobre la influencia de determinados alimentos y nutrientes específicos en este sentido.
La regulación de la ingesta de alimentos constituye un proceso esencial ya que posibilita el equilibrio necesario entre la cantidad de energía almacenada en forma de grasa corporal y el catabolismo de la misma. Este equilibrio es el resultado de la coordinación entre diferentes sistemas que van desde estructuras nerviosas centrales hasta los adipocitos (2). Los primeros estudios sobre la ingesta de alimentos mencionaban que el concepto de regulación de la ingesta se debía a una bajada de la glucosa y de los lípidos en la sangre como el principal elemento propiciador de la sensación de hambre. Actualmente se sabe que en la regulación de la ingesta participan numerosos péptidos, con acciones sinérgicas o antagónicas, que son sintetizados en diferentes tejidos (3).
El hipotálamo juega un papel importante en el control del apetito. En él se analizan las señales del intestino y del tronco cerebral y se procesan las señales para el control de la ingesta de alimentos. El hipotálamo también regula los ritmos circadianos: el núcleo supraquiasmático responde a la luz y sincroniza los ritmos conductuales y fisiológicos a través de oscilaciones circadianas de los circuitos cerebrales externos al núcleo supraquiasmático y en los tejidos periféricos, generando un ritmo de 24 horas en el comportamiento de la alimentación y el metabolismo (4).
La leptina es una hormona de 146 aminoácidos sintetizada por los adipocitos. Presenta efectos inhibitorios sobre la ingesta de alimentos ya que estimula el centro de la saciedad al unirse a sus receptores específicos hipotalámicos, disminuyendo la síntesis y liberación de diferentes neuropéptidos. Además, la secreción de leptina varía de acuerdo con el ritmo circadiano, incrementándose a lo largo del día y bajando durante el sueño nocturno. Por otra parte, es una hormona que favorece el gasto energético (5).
La grelina es un péptido compuesto por 28 aminoácidos secretada fundamentalmente por la mucosa del fondo gástrico. Es la única hormona orexígena periférica que activa los receptores localizados en los centros del apetito en el hipotálamo. La grelina es conocida como la "hormona del hambre" y es un péptido estimulador del apetito. En los seres humanos se produce un aumento preprandial y una caída posprandial de los niveles plasmáticos de grelina, lo que sugiere que el péptido desempeña un papel fisiológico en el inicio de la comida (6).
Los productos lácteos constituyen un importante grupo de alimentos y son una buena fuente de nutrientes en la dieta (7). En la actualidad, el consumo de productos lácteos ha disminuido por su posible implicación en el riesgo de padecer obesidad y trastornos metabólicos. Sin embargo, varios estudios observacionales y transversales han revelado una asociación inversa entre el consumo de productos lácteos y la composición corporal (8,9), aunque muchos aspectos de la fisiología de la regulación del apetito durante el consumo de productos lácteos aún no se conocen bien. Por tanto, con el presente estudio se pretende conocer cómo afecta el consumo de productos lácteos fermentados a la composición corporal y a la regulación del apetito en animales adultos sanos alimentados durante 30 días con dietas basadas en leche fermentada de vaca o de cabra.
MATERIAL Y MÉTODOS
DISEÑO EXPERIMENTAL Y DIETAS
Se han utilizado 20 ratas macho adultas (Ratus norvegicus, raza Wistar albina) procedentes del Servicio de Animales de Laboratorio de la Universidad de Granada. Los protocolos de manejo, cuidado y sacrifico de los animales empleados fueron aprobados por el Comité de Ética de la Universidad de Granada de acuerdo con las directrices comunitarias de la Unión Europea.
Las ratas fueron divididas en dos grupos experimentales (de 10 ratas cada uno), alimentados durante 30 días con dietas basadas en leche fermentada de vaca (raza Holstein) o leche fermentada de cabra (raza murciano-granadina). La composición de las dietas se encuentra en la tabla I. Se controló la ingesta de la dieta (pair feed) y los animales ingirieron agua bidestilada ad libitum.
Tabla I. Composición de las dietas experimentales (g/kg de dieta)
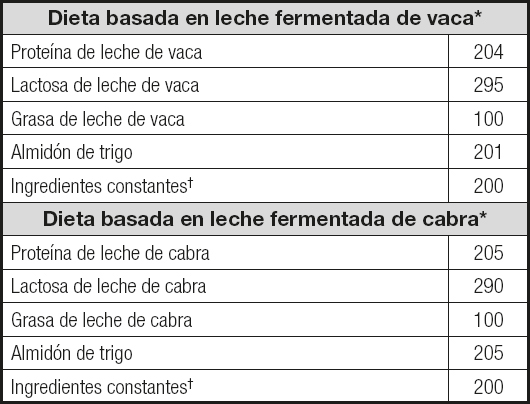
*Las dietas se prepararon de acuerdo con las recomendaciones del Instituto Americano de Nutrición al igual que los correctores minerales específicos, teniendo en cuenta el contenido mineral que aportaban las leches fermentadas (10).
†Los ingredientes constantes (g/kg de dieta) fueron: fibra (celulosa micronizada), 50 g; sacarosa, 100 g; cloruro de colina, 2,5 g; L-cistina, 3 g; corrector mineral, 35 g; y corrector vitamínico, 10 g
Al final del periodo experimental los animales se anestesiaron por vía intraperitoneal con pentobarbital sódico (Sigma-Aldrich Co., St. Louis, MO, EE. UU.) y se les extrajo sangre mediante canulación de la aorta abdominal. Se centrifugaron alícuotas de sangre con EDTA como anticoagulante para la obtención de plasma y posterior determinación de adipoquinas (leptina y adiponectina), hormonas reguladoras del metabolismo intermediario (grelina e insulina, hormona estimulante de la glándula tiroides, triyodotironina, tiroxina) y ácidos grasos no esterificados (AGNE).
FERMENTACIÓN Y DESHIDRATACIÓN DE LAS LECHES
Las leches fermentadas se prepararon según el método descrito previamente por Moreno-Fernández y cols. (11). Ambos tipos de leche se inocularon con iniciadores de yogur tradicional -Lactobacillus bulgaricus, subsp. Delbruickii, y Streptococcus thermophilus(concentración inicial de 1 x 1011UFC/ml de inóculo)- y se incubaron a 37 ºC aproximadamente. Previamente al proceso de deshidratación, la muestra de yogur se acondicionó y trató adecuadamente, y se homogeneizó calentándola a 20 ºC y agitándola vigorosamente con vórtex. Una vez preparado, el producto se deshidrató a una temperatura suave (50 ºC) para evitar cambios nutricionales negativos hasta que la humedad final se encontró entre el 2.5% y el 4.5%.
EVALUACIÓN DE LA COMPOSICIÓN CORPORAL
La composición del organismo completo (% de grasa y % de masa magra) se determinó usando la resonancia magnética cuantitativa (QMR) con un sistema Echo MRI Analyzer de Echo Medical Systems (Houston, Texas, EE. UU.). Todas las mediciones de la QMR se realizaron durante la fase de luz (09:00 a.m. a 06:00 p.m.). Los animales se colocaron en un cilindro de plástico de paredes delgadas (3 mm de grosor, 6,5 cm de diámetro interior) para limitar el movimiento. Posteriormente, los animales se sometieron brevemente a un campo electromagnético de baja intensidad (0,05 teslas) para medir la grasa, la masa magra, el agua libre y el agua corporal total. Este sistema genera una señal que modifica los patrones de giro de los átomos de hidrógeno dentro del sujeto experimental y utiliza un algoritmo para evaluar los cuatro componentes medidos. Las exploraciones con QMR se realizaron con tiempos de acumulación de 2 minutos.
DETERMINACIÓN DE HORMONAS TIROIDEAS, GRELINA, LEPTINA, ADIPONECTINA E INSULINA
La hormona estimulante del tiroides (TSH), la triyodotironina (T3) y la tiroxina (T4) se determinaron mediante el uso del panel de esferas magnéticas RYYMAG-30K Milliplex MAP Rat. La grelina (activa), la leptina y la insulina se determinaron usando el panel de esferas magnéticas de hormonas metabólicas de rata Milliplex MAP de RMHMAG-84K. Los niveles de adiponectina se midieron usando el ensayo de metabolismo del panel de adipocitos de rata Milliplex MAP RADPCMAG-82K (Millipore Corporation, Missouri, EE. UU.), basado en inmunoensayos en la superficie de microesferas magenéticas fluorescentes, siguiendo las especificaciones del fabricante (50 eventos por medida; 50 µl de muestra; configuración de ensayo: 8000-15.000; tiempo de espera, 60 segundos). La placa se leyó en el analizador LABScan 100 (Luminex Corporation, Texas, EE. UU.) con el software xPONENT para adquisición de datos. Los valores promedio de cada conjunto de muestras o estándares duplicados se encontraban dentro del 15% de la media. Las concentraciones de hormonas tiroideas, grelina, leptina, insulina y adiponectina en las muestras de plasma se determinaron comparando la media de las muestras duplicadas con la curva estándar de cada ensayo.
DETERMINACIÓN DE ÁCIDOS GRASOS NO ESTERIFICADOS
Los ácidos grasos no esterificados (AGNE) son moléculas liberadas por los triglicéridos por la acción de la enzima lipasa; se transportan en sangre unidos a la albúmina. Aportan solo una pequeña proporción de la grasa corporal; sin embargo, proporcionan una gran parte de la energía metabólica. Los AGNE se midieron en plasma usando un kit comercial (Randox Laboratories Ltd., Crumlin, Reino Unido).
ANÁLISIS ESTADÍSTICO
Los datos se muestran como medias ± error estándar de la media (SEM). El análisis estadístico se realizó con el programa informático SPSS (versión 22.0, 2013, SPSS Inc., Chicago, Illinois, EE. UU.). Las diferencias entre el grupo alimentado con dieta basada en leche fermentada de vaca y el alimentado con dieta basada en leche fermentada de cabra se evaluaron con la prueba de la t de Student. El nivel de significación se estableció en p < 0,05.
RESULTADOS
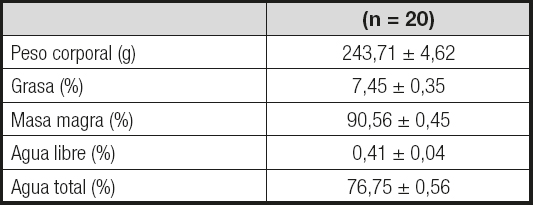
La composición corporal de los animales al inicio del estudio se detalla en la Tabla II. El peso corporal y los porcentajes de grasa, masa magra, agua libre y agua total se encontraban dentro del rango de normalidad para la edad de los animales estudiados.
Después del suministro durante 30 días de las dietas basadas en leche fermentada de vaca o cabra, los parámetros de composición corporal mostraron diferencias significativas (Tabla III). El peso y el porcentaje de grasa corporal fueron mayores en los animales alimentados con la dieta basada en leche fermentada de vaca (p < 0,001). Por el contrario, los porcentajes de masa magra, agua libre y agua total fueron mayores en los animales alimentados con la dieta basada en leche fermentada de cabra (p < 0,01 para la masa magra y p < 0,001 para el agua libre y total).
Tabla III. Composición corporal de las ratas alimentadas durante 30 días con dietas basadas en leche fermentada de vaca o cabra
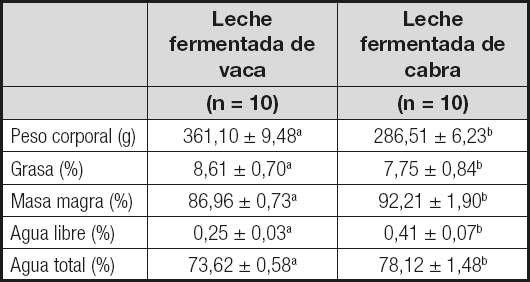
aLos valores de una misma fila con distinto superíndice son estadísticamente significativos (p < 0,05, test de la t de Student).
En la Tabla IVse presentan los parámetros plasmáticos relacionados con la regulación endocrina del metabolismo basal y la ingesta de alimento. No se registraron diferencias significativas en las concentraciones plasmáticas de hormonas tiroideas e insulina. Sin embargo, las concentraciones plasmáticas de grelina y adiponectina disminuyeron (p < 0,001), y los niveles de leptina y AGNE aumentaron (p < 0,001) en los animales alimentados con la dieta basada en leche fermentada de cabra en comparación con los alimentados con la dieta elaborada con leche fermentada de vaca.
Tabla IV. Concentración plasmática de hormonas que afectan al metabolismo basal y la ingesta de alimentos y ácidos grasos no esterificados en ratas alimentadas durante 30 días con dietas basadas en leche fermentada de vaca o cabra

TSH: hormona estimulante del tiroides; T3: triyodotironina; T4: tiroxina; AGNE: ácidos grasos no esterificados.
a,bLos valores de una misma fila con distinto superíndice son estadísticamente significativos (p < 0,05, test de la t de Student).
DISCUSIÓN
El tejido adiposo marrón es el principal tejido implicado en la termogénesis adaptativa de los pequeños mamíferos y los lactantes a través de la producción de calor mediante el desacoplamiento fisiológico de las mitocondrias. El desacoplamiento mitocondrial en el tejido adiposo permite la aceleración de la oxidación del sustrato sin que aumente en paralelo el ATP, de manera que la energía se pierde en forma de calor, proceso que es estimulado por las hormonas tiroideas (1,12). Por otro lado, el metabolismo también está regulado por el hipotálamo a través de un complejo circuito neuronal que controla la cantidad de energía que se ingiere y utiliza en cada momento. Los principales componentes que intervienen en esta regulación son la sensación de hambre y de saciedad, y la actividad del sistema nervioso autónomo y el sistema endocrino.
La realización de la mayoría de los procesos celulares inherentes a la vida depende de la disponibilidad de sustratos que garanticen un suministro energético adecuado. En reposo, la termogénesis adaptativa está controlada en gran medida por el sistema nervioso simpático, como ocurre durante la exposición al frío o la ingesta calórica excesiva. En ambas condiciones, el hipotálamo activa al sistema nervioso simpático para aumentar la producción de calor (1,13).
Los resultados del presente estudio concuerdan con otros que han demostrado los efectos beneficiosos del consumo de productos lácteos en la composición corporal (14,15). Sin embargo, estudios realizados con productos lácteos de leche de vaca (16) mostraron que la ingesta total de lácteos, yogur y proteína láctea estaba inversamente asociada a la grasa corporal; además, la ingesta de yogur y proteína láctea estaba inversamente asociada a la grasa abdominal. Estos autores también mostraron asociaciones negativas entre la ingesta de calcio y las medidas de adiposidad, lo que respalda aun más las investigaciones previas que muestran los beneficios de la suplementación con calcio sobre el peso corporal (17). Estudios anteriores han demostrado que el calcio puede aumentar la excreción fecal de ácidos grasos, incluidos las grasas saturadas y los ácidos biliares, minimizando los efectos sobre el colesterol sérico y aumentando la pérdida de energía, lo que puede afectar a las medidas de adiposidad a través de la lipogénesis y la lipólisis (18).En este sentido, la leche de cabra aumenta la biodisponibilidad del calcio (19) debido al mayor contenido de vitamina D, que favorece el transporte transcelular saturable de calcio dependiente de energía, por lo que podría contribuir a justificar el efecto de la leche fermentada de cabra en la disminución de la adiposidad.
Los productos lácteos contienen una amplia gama de nutrientes que incluyen proteínas (suero y caseína), aminoácidos de cadena ramificada, y péptidos. En este sentido, la leche de cabra y sus derivados lácteos contienen una cantidad algo menor de caseínas y una mayor proporción de proteínas séricas, lo que explica la mejor utilización digestiva de la proteína de la leche de cabra con respecto a la de vaca (20,21)..Murphy y cols., en 2013 (16), también mostraron una relación inversa entre las proteínas lácteas y todas las medidas de adiposidad. Estos resultados refuerzan las investigaciones previas que indican que la proteína láctea podría ser el componente responsable de los efectos beneficiosos sobre la composición corporal (22).
Estos efectos potenciales de la leche fermentada de cabra podrían estar relacionados con la influencia en el metabolismo lipídico de los adipocitos y, específicamente, con un aumento de la termogénesis inducida por la dieta, lo que conduciría a un mayor gasto de energía y a un menor almacenamiento de grasa. Otro posible mecanismo asociado al consumo de leche fermentada de cabra y la mejora de la composición corporal podría estar relacionado con la evidencia actual que respalda el consumo de proteína láctea y el papel de los lácteos en la saciedad, favoreciendo la pérdida de peso y la prevención del aumento de peso (23). En este sentido, la leche fermentada de cabra induce la elevación de los niveles plasmáticos de leptina y la reducción de los de grelina, disminuyendo el apetito, aumentando la sensación de saciedad y, por consiguiente, reduciendo el peso corporal.
En el presente estudio, las concentraciones plasmáticas de adiponectina disminuyeron en los animales alimentados con leche fermentada de cabra. Los AGNE mostraron una correlación inversa con las concentraciones plasmáticas de adiponectina, resultados que están de acuerdo con Kabara y cols. (24), quienes concluyeron que los niveles plasmáticos de adiponectina se correlacionan inversamente con las concentraciones plasmáticas de AGNE, un marcador importante de la movilización de lípidos. En este sentido, las menores concentraciones de adiponectina observadas en las ratas alimentadas con leche fermentada de cabra pueden conducir a un aumento de las tasas de lipólisis en el tejido adiposo. Utilizando ratones knockoutdel gen de adiponectina, Qiao y cols. (25) demostraron que esta adipoquina reduce la actividad de la lipasa sensible a hormonas al disminuir la estabilidad de la proteína-quinasa A. Por lo tanto, las reducciones fisiológicas de los niveles de adiponectina tras el consumo de leche fermentada de cabra pueden mejorar la lipólisis en el tejido adiposo. En el presente estudio, la estimulación de la lipólisis en el tejido adiposo se evaluó mediante el aumento de las concentraciones de AGNE y reducciones significativas en la masa de tejido adiposo visceral. El tejido adiposo visceral, debido a su ubicación anatómica y su drenaje portal, proporciona un acceso directo al hígado a los AGNE y las adipoquinas derivadas del tejido adiposo, en contraste con el tejido adiposo subcutáneo (26). Además, el tejido adiposo visceral tiene una mayor concentración de receptores beta-adrenérgicos y una mayor actividad de las enzimas lipolíticas, incluida la lipasa sensible a hormonas (27). Durante los periodos de intensa movilización de lípidos, la grasa adiposa visceral podría ser más sensible a los estímulos lipolíticos y la mayor liberación de AGNE podría reducir aun más la expresión de adiponectina (24). Por lo tanto, la conexión de la lipólisis adiposa visceral con los niveles de adiponectina podría ser un factor importante que influiría en la composición corporal.
CONCLUSIONES
Los resultados del presente estudio revelan que la dieta basada en leche fermentada de cabra influye en el metabolismo lipídico, disminuyendo la adiposidad e incrementando el gasto energético. Además, el consumo habitual de leche fermentada de cabra induce una elevación de la concentración plasmática de leptina y una reducción de la de grelina, disminuyendo el apetito y aumentando la sensación de saciedad y, por consiguiente, reduciendo el peso corporal. Finalmente, el consumo de la dieta basada en leche fermentada de cabra reduce la concentración plasmática de adiponectina, lo que conduce a un aumento de la lipólisis en el tejido adiposo y a un incremento de la concentración de ácidos grasos libres no esterificados, reduciendo la masa de tejido adiposo visceral.

