










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 texto en
texto en  Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkArchivos de la Sociedad Española de Oftalmología
versión impresa ISSN 0365-6691
Arch Soc Esp Oftalmol vol.80 no.12 dic. 2005
ARTÍCULO ORIGINAL
TOPOGRAFÍA DE LA SUMACIÓN ESPACIAL EN EL CAMPO VISUAL CENTRAL
SPATIAL SUMMATION: ITS TOPOGRAPHY IN THE CENTRAL VISUAL FIELD
GONZÁLEZ-HERNÁNDEZ M1, GONZÁLEZ-DE-LA-ROSA M1,
PAREJA A1, LOZANO V2, MESA F2
| RESUMEN Objetivo: La luminancia del estímulo (L) y su área (A) están relacionados por la ecuación: LxAk=constante. Se evaluó el valor de k en 66 posiciones del campo visual central. Palabras clave: Perimetría, campo visual, sumación espacial, campo receptivo, umbral | ABSTRACT Purpose: Luminance (L) and area (A) of the stimulus are related by the equation LxAk=constant. We evaluated the k value in 66 positions of the central visual field. Key words: Perimetry, visual field, spatial summation, receptive field, threshold. |
Recibido: 24/10/05. Aceptado: 12/12/05.
Hospital Universitario de Canarias. Universidad de La Laguna. Islas Canarias. España.
1 Doctor en Medicina.
2 Licenciado en Medicina.
Presentado al XVI Congreso de la Sociedad Internacional de Perimetría (IPS) (Barcelona 2004).
El Dr. Manuel González-de-la-Rosa tiene interés financiero en la estrategia perimétrica TOP y en el perímetro PULSAR usados en este trabajo.
Correspondencia:
Marta González-Hernández
C/. 25 de Julio, 34
38004 Santa Cruz de Tenerife
España
E-mail: martagh@jet.es
INTRODUCCIÓN
Desde los trabajos de Goldmann (1) usando la perimetría cinética clásica se aceptaba que podía obtenerse la misma isóptera con diversas combinaciones entre tamaño de estímulo e intensidad luminosa. Estas equivalencias han sido definidas por la ecuación LxAk=constante, donde L es la luminancia del estímulo y A su área. La escala de tamaños de Goldmann es universalmente conocida hoy en día; el diámetro del estímulo se duplica para cada paso, desde el nivel 0 (0,054º). De esta manera, por ejemplo el tamaño de Goldmann III equivale a un estímulo circular con un diámetro de 0,431º.
Si L1 y L2 son los valores de luminancia que definen el umbral para unos estímulos de área A1 y A2, el valor de k se obtendría mediante la siguiente fórmula: k = (Log L1 - Log L2) / (Log A2 - Log A1). Para que se cumplan las equivalencias del perímetro de Goldmann el valor de k sería aproximadamente 0,83 (2).
Desde que se inició el proceso de definir una perimetría con rigor fotométrico, diversos autores han estimado el valor de k en algunas regiones del campo visual utilizando tanto perimetría estática (3,4) como cinética (5) utilizando el perímetro manual de Goldmann. Una vez desarrollada la perimetría automática, el umbral luminoso empezó a medirse en decibelios (dB) en relación a la luminancia máxima del instrumento [log10(Lmax/L)]. En este momento Fankhauser (6) estimó una diferencia entre los tamaños Goldmann I y II de entre 3 y 4 dB en la fóvea (lo que equivale a k=0,291) y de 12 dB a 50 grados de excentricidad (k=0,998). Gramer (7) estimó un cambio entre los tamaños de Goldmann I y III así como entre los tamaños III y V de 6-10 dB, lo que equivaldría a un valor de k=0.499-0.832. Garway-Heath (8) usando cinco tamaños (Goldmann I-V) en el perímetro Humphrey calculó un valor de k de, aproximadamente, 0,33 en la fóvea y 0,63 a 30 grados de excentricidad.
Por otra parte desde los primeros estudios que hemos señalado, se indicó que la constancia definida por la fórmula LxAk=constante no era más que una aproximación, definida para unas áreas de estímulo centradas en el tamaño III de Goldmann. El valor de k se reduce para tamaños superiores y, por el contrario, se aproxima a la unidad para áreas muy pequeñas. Por lo tanto, la relación entre Log L y Log A es curvilínea y podría estimarse un valor diferente de k calculando la pendiente de la recta que es tangente a esta curva en cada valor de A (8).
Nuestra intención ha sido calcular el valor de k en 66 posiciones del campo visual central para observar si existen diferencias topográficas, o si puede admitirse un incremento progresivo del valor de k desde el centro a la periferia en cualquier meridiano, así como conocer la dependencia de este factor respecto al tamaño del estímulo.
SUJETOS, MATERIAL Y MÉTODO
Se utilizó el perímetro PULSAR (Prototipo experimental Octopus. HAAG-STREIT, Gartenstadtstrasse 10 CH-3098 Köniz. Berna. Suiza) provisto de un fotómetro digital que se sitúa en una esquina de un monitor Sansung SyncMaster 959NF (Sansung Electronics Co, 416 Maetan-3Dong, Paldal-Gu, Suwon City, Kyungki-Do, Corea) de manera que informa de la luminancia a un computador vía USB. De esta manera el computador regula periódicamente y de forma automática el brillo y el contraste para alcanzar la escala de luminancias deseada y mantenerla estable, pudiéndose seleccionar los 256 niveles de estímulo blanco habituales en los programas informáticos convencionales. La variación máxima de luminancia entre las diversas zonas de la pantalla se situó en el orden del 1%. La pantalla fue ajustada para una temperatura de color de 6500º K y verificada por medio de un colorímetro Tektronix J17 LumaColor.
Seleccionando un nivel de fondo de 31.5 asb pueden generarse estímulos diferenciales equivalentes a los 13-34 dB de un perímetro Octopus serie 300. La utilización de una resolución de 1024x768 píxeles en el monitor permite modificar el número de píxeles del estímulo para seleccionar cualquier tamaño entero o fracción en la escala Goldmann o modificar el tamaño del estímulo con la excentricidad, para compensar los cambios de distancia que se producen en una pantalla tangente. La necesidad de utilizar tamaños Goldmann fraccionarios nos ha obligado a prescindir de la clásica numeración romana que suele utilizarse habitualmente para definirlos. La limitación de rango dinámico que hemos señalado no significa que existan dificultades para el estudio de sujetos normales, siempre que el umbral que se pretenda medir no se sitúe por debajo de los 13 dB ni por encima de 34 dB.
La disposición de los puntos de examen fue equivalente a la del programa 30-2 de Octopus, excluyendo las filas superior e inferior (horizontal= -30º a +30º) x (vertical= +24º a -24º), con una separación entre estímulos de 6 grados. Al usarse un monitor de 19 pulgadas la distancia del ojo del paciente al punto de fijación, situado en el centro de la pantalla, fue de 31cm. En los pacientes présbitas se utilizó la graduación de cerca correspondiente. Los falsos positivos y negativos fueron examinados de manera tradicional, es decir falsos positivos en el caso de respuesta en ausencia de estímulo y falsos negativos en el caso de ausencia de respuesta a un estímulo con intensidad por encima del umbral previamente determinado. Se interpretaron como errores de fijación los casos de respuesta a estímulos proyectados sobre la mancha ciega (método de Heijl-Krakau).
Se estudió el umbral luminoso en 66 puntos del campo visual central usando la estrategia TOP (9) para estímulos de cinco tamaños diferentes (Goldmann 1,9, 2,5, 3, 3,5 y 4), en 10 ojos de 10 sujetos sanos.
La presentación del estímulo fue de 200 mseg, que es el tiempo de presentación usado por los perímetros Humphrey para evitar la interferencia del parpadeo en la detección del estímulo. El perímetro Octopus usa un tiempo de presentación de 100 mseg., ya que usa una cámara de vídeo para eliminar aquellos estímulos que coinciden con el parpadeo.
Todos los exámenes fueron realizados en la misma sesión, con un mínimo de 10 minutos de descanso entre cada uno de ellos, manteniendo al paciente en un ambiente de adaptación fotópica baja. El orden de los exámenes fue aleatorio. Inmediatamente antes de cada examen se realizó una verificación fotométrica automática del instrumento.
Todos los sujetos eran voluntarios con amplia experiencia perimétrica, con conocida capacidad de atención y motivación en las pruebas, sin patología ocular ni general o tratamientos que pudiesen alterar los resultados perimétricos. Todos los sujetos tenían agudeza visual unidad o superior, ametropía inferior a 2 dioptrías (D) de esfera y 1,75 D de cilindro. Pese a la experiencia perimétrica previa se realizó un período de adaptación a la prueba y se verificó que los resultados contenían menos de un 25% de errores (falsos positivos y negativos y errores de fijación).
Se calculó el resultado de la ecuación LxAk para todos los valores de k comprendidos entre 0.001 y 1 en pasos de 0,001 (1.000 niveles) utilizando el valor de luminancia correspondiente al umbral medio obtenido en cada una de las 66 posiciones examinadas para los 5 tamaños de estímulo. A continuación se calculó el coeficiente de variación de Pearson de los cinco valores resultantes en cada uno de los 1.000 niveles y se definió como valor de k de cada posición aquel nivel en el que el coeficiente de variación de Pearson alcanzó su valor mínimo. La figura 1 muestra un ejemplo de este cálculo aplicado a los valores medios de sensibilidad media (MS) de la población estudiada.
RESULTADOS
La media de edad de los sujetos examinados fue de 38,5 años (d.e.= 16,6). Seis eran del sexo femenino y cuatro del masculino. La duración media de los exámenes fue de 2:57 minutos (d.e.= 0:12) y el número medio de errores 3,1%.
Los valores de umbral obtenidos con el tamaño menor (Goldmann 1.9) presentaron una media de 19.4 dB (d.e.=3,8) y los del tamaño mayor (Goldmann 4) un valor medio de 27,2 dB (d.e.=2,6) sin que se superasen los límites de rango dinámico del instrumento, por sus extremos inferior o superior.
En comparación con la sensibilidad obtenida para el tamaño 4, la sensibilidad se redujo 1,5, 2,9, 5,4 y 7,8 dB para los tamaños siguientes. Por lo tanto se calculó una media de k=0,616 para todo el campo visual (fig. 1).
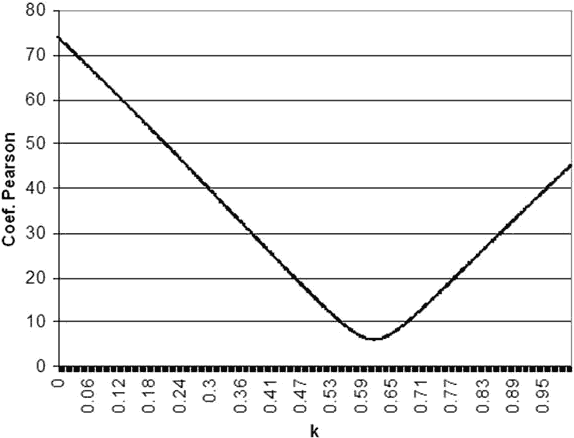
Fig. 1. Ejemplo del cálculo de k usando valores medios de umbral (MS medio) en la población examinada.
El umbral medio obtenido con los tamaños de estímulo 1,9, 2,5, 3, 3,5 y 4 (áreas 0,854, 1,993, 4,271, 8,447 y 16,040 mm2)
fueron 19,4, 21,8, 24,3, 25,7 y 27,2 dB. Las intensidades luminosas correspondientes con esos niveles fueron 46,3, 26,7, 14,9, 10,9 y 7,7 asb.
El coeficiente de variación de Pearson se aplicó a los cinco valores resultantes de la ecuación LxAk,
es decir: 46,3x(0,854k), 26,7x(1,993k), 14,9x(4,27k), 10,9x(8,447k) y 7,7x(16,040k), para valores de k entre 0 y 1.
El coeficiente de Pearson más bajo (mejor constancia entre las cinco ecuaciones) fue 6,0, correspondiendo a un valor de k de 0,616.
La figura 2 muestra un ejemplo del cálculo de k en tres puntos del campo visual siguiendo el procedimiento descrito. La figura 3 indica los valores de k estimados en los 66 puntos del campo visual. Se puede observar que el valor de k en el campo visual central no es muy diferente del estimado en algunas áreas supero-nasales periféricas, aunque los umbrales sean muy diferentes.
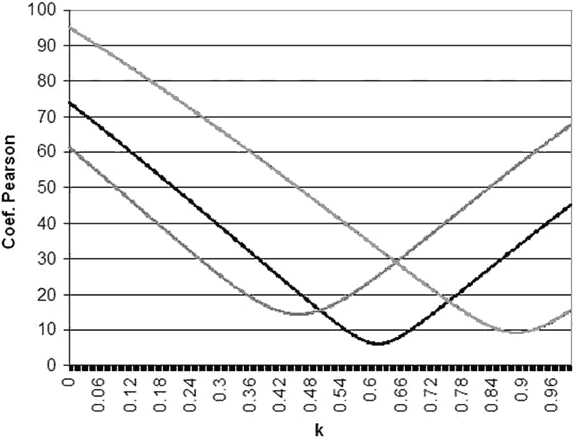
Fig. 2. Cálculo de k en tres puntos del campo visual: Izquierda. Punto supero-nasal (X=-21, Y=15), k=0,46.
Centro. Punto supero-temporal (X=27, Y=3), k=0,64. Derecha. Punto infero-nasal (X=-21, Y=-15), k=0,89.
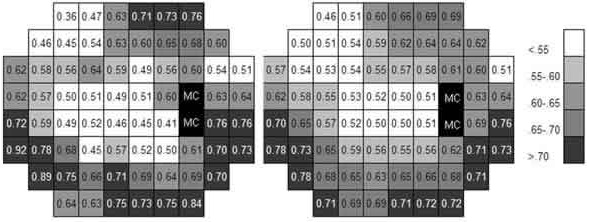
Fig. 3. La imagen izquierda muestra los valores de k calculados en cada uno de los 66 puntos examinados.
En la imagen derecha se muestran los valores medios de cada área (media de cada punto y los que le rodean)
para suavizar las diferencias locales, probablemente producidas por error.
El esquema corresponde al ojo derecho. "MC" corresponde a la mancha ciega.
La tabla I muestra los valores medios de los umbrales obtenidos en los cuatro cuadrantes del campo visual y el valor medio de k. Esta constante presenta su valor menor en el cuadrante supero-nasal y el mayor en el cuadrante infero-nasal.

El valor de k aumenta de manera lineal desde el centro hacia la periferia pero con pendientes desiguales en los diversos meridianos. Cuando se calcula la relación entre k y la excentricidad de los 66 puntos se obtiene un coeficiente de correlación relativamente bajo (r= 0,52, p<0,01) debido al comportamiento diferente de k en cada meridiano. Sin embargo, cuando los puntos se agrupan en sus 10 valores de excentricidad, las medias de k alcanzan una alta correlación con la excentricidad (r=0.98, p<0,01) con una pendiente de 0,01 por grado (fig. 4).
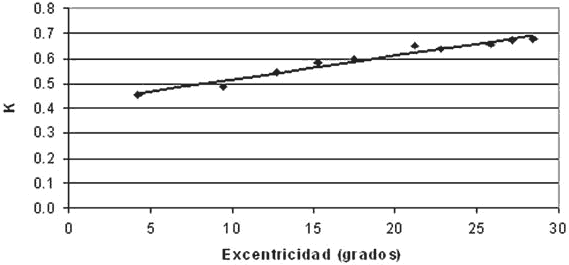
Fig. 4. Promedio de los valores de k en 10 niveles de excentricidad en los que se agruparon los 66 puntos estudiados.
Cuando se analizan por separado los resultados obtenidos con los tamaños 1,9, 2 y 3 de Goldmann los valores de k se incrementan respecto a los estimados usando los cinco tamaños (fig. 5). Por el contrario cuando se realiza un análisis teniendo en cuenta solamente los resultados obtenidos con los tamaños 3, 3,5 y 4 el valor de k se reduce. Sin embargo las características que se describieron previamente para las diferentes áreas del campo visual, se mantienen de forma aproximada, siendo el cuadrante nasal superior el que presenta valores inferiores de k, resultando similares a los que se obtienen en la región paracentral.
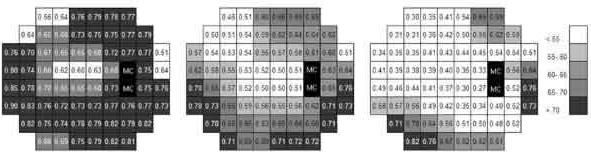
Fig. 5. Valores de k obtenidos usando tamaños de Goldmann 1,9, 2 y 3 (izquierda)
y tamaños 3, 3,5 y 4 (derecha) en comparación con los valores medios de k (centro).
DISCUSIÓN
Los valores medios de k que hemos obtenido son comparables a los descritos por otros autores utilizando perímetros comerciales y procedimientos matemáticos diferentes. De la misma manera los valores medios de incremento de k con la excentricidad son similares a los que se muestran o se deducen de los resultados de otros trabajos (3,7,10-13). Nuestro procedimiento difiere esencialmente en la estrategia de estudio del campo visual, que se adapta a las características locales de dependencia entre los puntos próximos, actuando como un promediador local, así como también en el procedimiento matemático de cálculo del valor de k. El análisis del coeficiente de variación de Pearson aplicado a los resultados experimentales, permite obtener resultados altamente congruentes, tal y como puede comprobarse al observar las gráficas expuestas en las figuras 1 y 2, aportando unas estimaciones locales detalladas de la sumación espacial en todo el campo visual central, que son la aportación principal del presente trabajo.
Se había señalado un comportamiento similar de la sumación espacial entre el meridiano vertical y el horizontal (10) y que las diferencias entre cuatro meridianos eran una cuestión de escala (2). Nuestros resultados indican, por el contrario, un comportamiento muy diferente de la sumación espacial en el campo visual superior respecto al inferior, posiblemente porque los meridianos en los que k varía más (entre120º y 150º para ojo derecho) no fueron analizados en estos estudios. Los cambios topográficos de los umbrales normales no parecen correlacionarse con los observados en la sumación espacial.
Si, como expone Galrway-Heath (8) el área del estímulo puede ser representada por el número de células ganglionares en la ecuación LxAk=constante, habría que suponer una desigual distribución de estas células que explicase el comportamiento asimétrico observado en la sumación espacial. Sin embargo varios estudios han informado que la retina inferior posee una menor concentración de células ganglionares que la superior (14,15), de manera que en el campo visual superior cabría esperar en todo caso un incremento del valor de k respecto al inferior, siendo así que en nuestra experiencia ocurre justamente lo contrario.
Se ha confirmado que el valor de k varía con el rango de tamaños de estímulo que se analice pero manteniéndose unas diferencias topográficas relativamente constantes. Por lo tanto las diferencias en sensibilidad luminosa y agudeza visual entre la región paracentral del campo visual y algunas zonas más periféricas del campo superior, que explican la magnificación cortical (16) no se acompañan aparentemente de un cambio proporcional en la sumación espacial. La sensibilidad diferencial a la luz y otros parámetros fisiológicos se degradan hacia la periferia en relativa dependencia con la concentración local de células ganglionares, pero en el caso del valor de k habría que admitir que este no puede ser el único factor que condiciona la sumación espacial. Debemos suponer que, como con otras funciones visuales, existen variaciones en las características fisiológicas de las células ganglionares a través de la retina (17) u otros factores que expliquen esta disparidad. Como ejemplo pueden señalarse las diferencias observadas en el diámetro axonal de las células ganglionares procedentes de las hemi-retinas superior e inferior (18), las diferencias contrapuestas en la concentración de células horizontales A y B entre ambas (19) o diversos estudios que informan de una mayor densidad de conos en la hemi-retina inferior (20,21).
BIBLIOGRAFÍA
1. Goldmann H. Grundlagen exakter Perimetrie. Ophthalmologica 1945; 109: 57-70. [ Links ]
2. Latham K, Whitaker D, Wild JM. Spatial summation of the differential light threshold as a function of visual field location and age. Ophthalmic Physiol Opt 1994; 14: 71-78. [ Links ]
3. Gougnard L. Étude des summations spatiales chez le sujet normal par la périmétrie statique. Ophthalmologica 1961; 142: 469-486. [ Links ]
4. Dannheim F, Drance SM. Studies of spatial summation of central retinal areas in normal people of all ages. Can J Ophthalmol 1971; 6: 311-319. [ Links ]
5. Verriest G, Ortiz-Olmedo A. Étude comparative du seuil différentiel de luminance et de l'exposant de sommation spatiale pour des objets pleins et pour des objets annulaires de mèmes surfaces. Vision Res 1969; 9: 267-282. [ Links ]
6. Fankhauser F. Problems related to the design of automatic perimeters. Doc Ophthalmol 1979; 47: 89-139. [ Links ]
7. Gramer E, Kontic D, Krieglstein GK. Die computerperimetrische Darstellung glaukomatoser Gesichtsfelddefekte in Abhangigkeit von der Stimulusgrosse. Ophthalmologica 1981; 183: 162-167. [ Links ]
8. Garway-Heath DF, Caprioli J, Fitzke FW, Hitchings RA. Scaling the hill of vision: the physiological relationship between light sensitivity and ganglion cell numbers. Invest Ophthalmol Vis Sci 2000; 41: 1774-1782. [ Links ]
9. Gonzalez de la Rosa M, Martinez A, Sánchez M, Mesa C, Cordoves L, Losada MJ. Accuracy of the Tendency-Oriented Perimetry with the Octopus 1-2-3 Perimeter. In: Wall M, Heijl A. Perimetry Update 1996/1997. Amsterdam: Kugler; 1997; 119-123. [ Links ]
10. Sloan LL. Area and luminance of test object as variables in examination of the visual field by projection perimetry. Vision Res 1961; 1: 121-138. [ Links ]
11. Wilson ME. Invariant features of spatial summation with changing locus in the visual field. J Physiol 1970; 207: 611-622. [ Links ]
12. Wood JM, Wild JM, Drasdo N, Crews SJ. Perimetric profiles and cortical representation. Ophthalmic Res 1986; 18: 301-308. [ Links ]
13. Kasai N, Takahashi G, Koyama N, Kitahara K. An analysis of spatial summation using a Humphrey field analyzer. In: Mills RP. Perimetry Update 1992/1993. Amsterdam: Kugler; 1992; 557-562. [ Links ]
14. Curcio CA, Allen KA. Topography of ganglion cells in human retina. J Comp Neurol 1990; 300: 5-25. [ Links ]
15. Danckert J, Goodale MA. Superior performance for visually guided pointing in the lower visual field. Exp Brain Res 2001; 137: 303-308. [ Links ]
16. Rovamo J, Virsu V. An estimation and application of the human cortical magnification factor. Exp Brain Res 1979; 37: 495-510. [ Links ]
17. Wild JM, Wood JM, Flanagan JG. Spatial summation and the cortical magnification of perimetric profiles. Ophthalmologica 1987; 195: 88-96. [ Links ]
18. Fitzgibbon T, Funke K. Retinal ganglion cell axon diameter spectrum of the cat: mean axon diameter varies according to retinal position. Vis Neurosci 1994; 11: 425-439. [ Links ]
19. Muller B, Peichl L. Horizontal cells in the cone-dominated tree shrew retina: morphology, photoreceptor contacts, and topographical distribution. J Neurosci 1993; 13: 3628-3646. [ Links ]
20. Muller B, Peichl L. Topography of cones and rods in the tree shrew retina. J Comp Neurol 1989; 282: 581-594. [ Links ]
21. Curcio CA, Sloan KR, Kalina RE, Hendrickson AE. Human photoreceptor topography. J Comp Neurol 1990; 292: 497-523. [ Links ]

