










My SciELO
 Custom services
Custom servicesServices on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAnales del Sistema Sanitario de Navarra
Print version ISSN 1137-6627
Anales Sis San Navarra vol.31 n.1 Pamplona Jan./Apr. 2008
REVISIONES
Regeneración de la superficie ocular: stem cells/células madre y técnicas reconstructivas
Regeneration of the ocular surface: stem cells and reconstructive techniques
A. Fernández1, J. Moreno1, F. Prósper2, M.García2, J. Echeveste3
1. Departamento de Oftalmología. Clínica Universitaria de Navarra. Pamplona.
2. Área de Terapia Celular y Hematología. Clínica Universitaria de Navarra. Pamplona.
3. Departamento de Anatomía Patológica. Clínica Universitaria de Navarra. Pamplona.
Dirección para correspondencia
RESUMEN
La córnea es un tejido transparente constituido microscópicamente por 5 capas bien diferenciadas. El epitelio corneal es esencial para la transparencia corneal y se encuentra en continua renovación a lo largo de la vida a partir de la población de células madre limbocorneales. La localización de estas células madre limbocorneales parece residir en las capas basales del epitelio limbocorneal, de vital importancia para mantener el microambiente de estas células madre limbocorneales, que depende de una variedad de factores intrínsecos y extrínsecos. La insuficiencia límbica se produce cuando ocurre una pérdida parcial o total de estas células madre limbocorneales. Este cuadro lleva a una opacificación corneal con la consiguiente pérdida de visión. En estos casos, el trasplante corneal supone únicamente un reemplazo temporal del epitelio corneal; es necesario llevar a cabo un tratamiento previo con trasplante de limbo autólogo o alogénico, que permita regenerar la población de células limbocorneales dañadas. Para disminuir el riesgo que supone el trasplante de limbo en el ojo donante, se han propuesto técnicas de cultivo de células limbocorneales a partir de pequeñas biopsias limbocorneales.
Palabras clave. Epitelio corneal. Células madre limbocorneales. Insuficiencia límbica. Cultivo celular.
ABSTRACT
The cornea is a transparent tissue microscopically constituted by 5 well differentiated layers. The corneal epithelium is essential for corneal transparency and is found in a state of constant renovation throughout life on the basis of the population of limbocorneal stem cells. The localisation of these limbocorneal stem cells seems to be in the basal layers of the limbocorneal epithelium, of vital importance for maintaining the micro-environment of these limbocorneal stem cells, which depend on a variety of intrinsic and extrinsic factors. Limbic insufficiency occurs when there is a partial or total loss of these limbocorneal stem cells. These clinical features lead to a corneal clouding with a resulting loss of vision. In these cases, corneal transplant only represents a temporary replacement of the corneal epithelium; it is necessary to carry out a prior treatment involving transplant of the autologous or allogeneic limbus, which enables regeneration of the population of damaged limbocorneal cells. To reduce the risk involved in the transplant of the limbus of the donor eye, techniques of cultivation of limbocorneal cells on the basis of small limbocorneal biopsies are proposed.
Key words. Corneal epithelium. Limbocorneal stem cells. Limbic deficiency. Cellular cultivation.
Anatomía de la córnea y del limbo esclerocorneal
La córnea es un tejido transparente que une la esclera opaca y el limbo esclerocorneal y tiene una función refractiva así como de barrera química y mecánica entre el ojo y el medio ambiente.
Microscópicamente, la córnea puede dividirse en 5 capas (Fig. 1): epitelio, capa de Bowman, estroma, membrana Descemet y endotelio.

El epitelio es estratificado escamoso no queratinizado, consta de 4 a 6 capas de células y representa el 10% del grosor corneal. Morfológicamente está dividido en 3 capas:
– Una capa única de células columnares basales que se unen mediante hemidesmosomas a la membrana basal epitelial. Representan la capa germinativa del epitelio ya que son las únicas células epiteliales que tienen capacidad de dividirse por mitosis.
– Dos o tres filas de células que tienen finas extensiones en forma de alas. En su periferia se unen por uniones tipo desmosoma y tipo GAP.
– Dos capas de células de superficie alargadas y delgadas unidas por puentes. Después de un tiempo de vida de algunos días las células superficiales se desprenden a la película lagrimal. Debido a su excelente capacidad de regeneración, el epitelio no se escarifica.
La capa de Bowman es una estructura acelular que representa la capa superficial del estroma. Está formada por fibras de colágeno organizadas al azar. Esta capa no se regenera cuando se daña.
El estroma ocupa alrededor del 90% del grosor de la córnea y está compuesto principalmente por capas de fibrillas de colágeno orientadas de forma regular. La separación entre las capas es mantenida por una sustancia fundamental de proteoglicanos con un sincitio de fibroblastos modificados (queratocitos) intercalados entre las capas. Los axones nerviosos y sus células de Schwann asociadas se encuentran en el tercio medio anterior del estroma.
El colágeno constituye aproximadamente el 71% del peso seco de la córnea y la macromolécula estructural que proporciona la transparencia tisular así como la resistencia mecánica a la PIO. El colágeno tipo 1 es el que predomina.
Los queratocitos ocupan el 3-5% del volumen estromal. Su función consiste en mantener las fibras de colágeno y la matriz extracelular mediante una actividad de síntesis constante.
La membrana de Descemet es la lámina basal gruesa segregada por el endotelio. Está compuesta por un fino enrejado de fibrillas de colágeno. Consta de 2 capas, una zona estriada anterior, y una zona no estriada posterior, que está producida por el endotelio a lo largo de la vida.
El endotelio corneal se deriva de la cresta neural y consta de una única capa de células hexagonales. Desempeña una función vital en el mantenimiento de la turgencia de la córnea. Con la edad, el número de células endoteliales desciende gradualmente por lo que, debido a que no pueden regenerarse, las células vecinas han de progresar para llenar el espacio.
En el nacimiento, la densidad celular oscila entre 3.500 y 4.000 células/mm2, mientras que la córnea del adulto tiene densidades entre 1.400 a 2.500 células/mm2. Cuando las células endoteliales están sometidas a estrés, especialmente si se pierden algunas células, las células restantes pierden su forma hexagonal y se vuelven irregulares en forma (pleomorfismo) y en tamaño. Estos cambios pueden ocurrir con la edad, tras traumatismos y con el uso prolongado de lentes de contacto.
Desde un punto de vista clínico, es importante destacar que el endotelio corneal es funcionalmente esencial para la córnea. Es una capa frágil cuya integridad y viabilidad deben ser garantizadas para asegurar el éxito de cualquier proceso intraocular.
Recientemente se ha estudiado la capacidad de autorrenovación de los precursores del endotelio corneal en conejos y se ha visto como el endotelio periférico contiene más precursores y tiene una mayor capacidad de autorregeneración que el endotelio central1.
Una de las propiedades del endotelio corneal es su impermeabilidad a las sustancias hidrosolubles. La ruptura de esta barrera supone la penetración de moléculas que pueden afectar al medio intraocular dañando los tejidos corneales.
La superficie ocular está compuesta por tres epitelios distintos: corneal, limbar y conjuntival. Son todos ellos epitelios estratificados, escamosos y no queratinizados procedentes de la superficie ectodérmica. Sin embargo, difieren en sus características y sus funciones. Estas diferencias están reflejadas en sus patrones únicos de expresión genética.
Epitelio conjuntival
El epitelio conjuntival es un epitelio estratificado escamoso no queratinizado al igual que el epitelio corneal pero contiene células caliciformes secretoras de mucina que contribuyen a mantener la capa lagrimal de la superficie ocular y que se encuentran intercaladas entre células epiteliales (Fig. 2).

Está muy bien vascularizado. Las diferencias entre los fenotipos epiteliales conjuntival y corneal se basa en la expresión de diferentes queratinas, mucinas y glicocalix. Las citoqueratinas características del epitelio no queratinizado estratificado (CK4 y 13), y simple (CK8 y 19), se expresan en las capas superficiales del epitelio conjuntival2. Sus células precursoras se localizan en el fórnix conjuntival y la unión mucocutánea3. Las células progenitoras del fórnix tienen gran capacidad proliferativa, mucho mayor que las células localizadas en las regiones bulbar y palpebral4. La unión mucocutánea de la conjuntiva palpebral en conejos parece ser la principal fuente de células TAC (Transient Amplifying Cells) en continua replicación. Estas células migran hacia el fórnix para regenerar las células maduras que se pierden de forma continua3. Las células madre tienden a localizarse en áreas de mayor grosor del epitelio así como en zonas de mayor pigmentación. Se ha demostrado que las células progenitoras del epitelio conjuntival son bipotenciales, es decir, son células precursoras tanto de células caliciformes como de células no-caliciformes5. La diferenciación de las células caliciformes, sin embargo, parece requerir un medio ambiente estromal más específico, como demuestran algunos estudios sobre el cultivo de células epiteliales conjuntivales de ratón sobre membrana amniótica6. Los epitelios corneal y conjuntival son continuos y componen la superficie ocular. Sin embargo, sus perfiles de expresión genética son bastante diferentes.
Epitelio limbar
El epitelio limbar es la zona transicional entre los epitelios corneal y conjuntival. Morfológicamente es diferente de la córnea en que posee células de Langerhans y melanocitos y de la conjuntiva en que carece de células caliciformes7. El epitelio limbar se encuentra sobre un estroma altamente vascularizado, de donde le llega el aporte sanguíneo. Los vasos sanguíneos forman parte de las empalizadas de Vogt (Fig. 3), que permiten una aproximación entre los vasos sanguíneos y el epitelio, proporcionándole altos niveles de nutrición y citoquinas. Estas citoquinas tienen un papel importante en el mantenimiento de las células madre limbares y tienen 3 patrones de expresión, las citoquinas tipo I, II y III8.

Ya ha sido demostrado cómo las células madre del epitelio corneal tienen una localización en la capa basal de la córnea periférica, en la zona limbar, jugando un papel fundamental en la proliferación y diferenciación celular del epitelio corneal9,10. Esta idea se apoya en varios hechos: el patrón único de expresión de la citoqueratina 3 en las células corneales diferenciadas, la localización de las células de ciclo lento en el limbo, el mayor potencial proliferativo de las células limbares en comparación con las células corneales centrales tanto in vivo como in vitro y la capacidad de las células limbares basales de regenerar un epitelio corneal dañado10. El epitelio corneal utiliza tres estrategias para expandir su población celular durante la cicatrización: el reclutamiento de células madre para producir más células TAC, el aumento de la tasa de replicación de las células TAC y el aumento de la eficacia de la replicación de las células TAC con el acortamiento del ciclo celular11. Las células madre limbocorneales se encuentran en la capa basal del epitelio limbar mientras que las células TAC se encuentran en la capa basal de todo el epitelio corneal. La localización limbar de las células madre limbocorneales explica propiedades corneales como el predominio de formaciones tumorales en la zona limbar y la migración centrípeta de las células corneales periféricas hacia la córnea central. Este movimiento celular centrípeto es el responsable de la regeneración del epitelio corneal central12,13. Conforme las células se mueven centrípetamente, van desarrollando características de células TAC y adquiriendo marcadores de novo, como las citoqueratinas 3/12. Schermer y col9, en cultivos de células corneales de conejo, observaron que la citoqueratina 3 se asociaba a las capas superiores y más diferenciadas, lo que implica que esta citoqueratina 3 constituye un marcador de estados avanzados de diferenciación. También se expresaba uniformemente en el epitelio corneal central de los conejos. Esta expresión homogénea sugiere que, aunque las células basales en la zona limbar son indiferenciadas, las del epitelio corneal central son más diferenciadas en función de la expresión de citoqueratina 3. Esto les llevó a proponer que las células madre del epitelio corneal no se distribuyen uniformemente a lo largo de la capa basal del epitelio corneal completo9. La preservación de las células madre limbocorneales parece depender de las condiciones ambientales. Su regeneración requiere unas condiciones específicas que sólo se encuentran en el nicho de localización de estas células madre. Fuera de estas condiciones, las células madre proliferan desarrollando una expresión gradual de diferentes marcadores celulares. Durante la diferenciación, se van expresando nuevos genes a la vez que se suprimen otros previamente expresados. Los estados intermedios presentan una gran capacidad proliferativa. Tras un número de replicaciones, esta capacidad va disminuyendo hasta alcanzar la diferenciación terminal. Llegado un momento, las células TAC cesan sus mitosis y entran en el compartimento no proliferativo, pasando a la diferenciación celular terminal14. El esquema de diferenciación seguido es de stem cells a células TAC y a células diferenciadas9,15. La capa basal del epitelio limbar contiene tanto células madre como células TAC, así como los precursores tempranos que pueden existir entre ambos tipos celulares. El epitelio corneal está constituido por una jerarquía de células TAC, aquellas localizadas en la periferia con capacidad de múltiples divisiones y las células TAC de la córnea central con capaces de dividirse tan sólo una vez11. El hecho de que las células TAC de la córnea central tengan una escasa capacidad de replicación concuerda con los estudios que muestran que las células de la córnea central constituyen colonias terminales que no pueden ser cultivadas más de dos veces16. La transición de células madre a células TAC no puede ser identificada con marcadores químicos. La localización anatómica de las células madre puede ser identificada por las empalizadas de Vogt17 (Fig. 3), que están pigmentadas, y se encuentran en el margen de la córnea clara.
Parece que las células madre limbocorneales no se encuentran en igual proporción en los distintos cuadrantes, siendo su proporción mayor en la córnea superior e inferior en comparación con los cuadrantes temporal y nasal18. El marcador molecular DNp63 está altamente expresado en el epitelio limbar. Otros marcadores negativos, como la conexina 43, también se usan para identificar las células basales del limbo más inmaduras. Una serie de rasgos anatómicos, como la presencia de melanocitos, sugieren la existencia de este nicho de células madre necesario para la supervivencia de estas células madre durante el ciclo celular. Parece que existe otra población celular (Side Population) en el epitelio limbar, que también expresa la proteína ABCG2, al igual que las células madre hematopoyéticas de la médula ósea19.
Células madre
Limbocorneales
Las células madre son una pequeña subpoblación del total de los tejidos, caracterizadas por una rápida y continua renovación celular. Las células diferenciadas son células con un ciclo de vida corto que se regeneran por medio de la proliferación de la subpoblación de células madre. Las células madre fueron definidas por Potten y Loeffler20 como células indiferenciadas capaces de proliferar, automantenerse, dar lugar a una gran progenie celular diferenciada, regenerar los tejidos tras un daño y poseer gran flexibilidad en cada una de estas capacidades13. Las características principales de las células madre son el estar pobremente diferenciadas con un citoplasma primitivo, el tener una alta capacidad de autorrenovación sin errores, una larga esperanza de vida, un ciclo celular largo y el ser capaces de llevar a cabo divisiones simétricas o asimétricas, activándose su proliferación por la cicatrización o por su puesta en cultivo. Por definición, las células madre están presentes en todos los tejidos autorrenovables con un alto grado de diferenciación celular. Estas células permanecen relativamente quiescentes, pero tienen un gran potencial para la división celular clonogénica y son responsables de la proliferación y la diferenciación celulares. Las células madre pueden llevar a cabo divisiones celulares asimétricas, es decir, originando una célula hija que permanece indiferenciada y otra célula hija destinada a la diferenciación y división celular, las células amplificadoras transitorias o células TAC. Estas células, por mitosis, aumentan el número de células, diferenciándose en células postmitóticas. Las células madre y las células TAC constituyen el compartimento proliferativo del tejido, diferenciándose en su ciclo vital y en la actividad mitótica. Las células TAC tienen un potencial proliferativo limitado, se dividen con más frecuencia y dan lugar a células diferenciadas incapaces de dividirse20. Las células madre de muchos tejidos están siendo estudiadas como tratamiento de enfermedades degenerativas intratables de ninguna otra forma21. Schermer y col9 determinaron la localización limbar de estas células madre limbocorneales a partir de un estudio sobre la expresión de la keratina corneal de 64kD. También sugirieron que las células TAC están localizadas en la capa basal corneal y las células postmitóticas en las capas suprabasales. Conforme las células madre limbocorneales van migrando desde el limbo al centro de la córnea, se van diferenciando en células epiteliales corneales maduras o diferenciadas. Se han estudiado diversos genes, como el gen de la involucrina humano22, que pueden estar implicados en la regulación de los cambios que ocurren durante esta diferenciación de células madre limbocorneales a células TAC. Actualmente, la localización limbar de las células madre del epitelio corneal se basa en varios datos:
– La ausencia de marcadores específicos del epitelio corneal diferenciado, como son las citoqueratinas 3 y 12.
– La presencia de células de ciclo lento en el epitelio limbar basal, con una gran capacidad proliferativa en cultivo.
– La cicatrización corneal anormal con conjuntivalización, vascularización e inflamación crónica en casos de daño parcial o completo del epitelio limbar.
– La utilización de las células limbares en la reconstrucción del epitelio corneal de pacientes con insuficiencia límbica23.
Existe una hipótesis propuesta por Thoft y col24 que explica la renovación del epitelio corneal por medio de tres ejes (Fig. 4): el X, que consiste en la proliferación de las células basales, el Y, que consiste en la proliferación y migración centrípeta de las células limbares y el Z, que consiste en la pérdida de células epiteliales de la superficie corneal.

El mantenimiento del epitelio corneal, por tanto, puede definirse por medio de la ecuación: X + Y = Z (Fig. 4), la cual epresenta cómo, para mantener el epitelio corneal, la pérdida celular debe estar en equilibrio con el reemplazo celular. El componente Y es un movimiento celular centrípeto que ocurre incluso en ausencia de un defecto agudo. Es importante no confundir Y con otro fenómeno, el movimiento rápido de las células periféricas en respuesta a un defecto central agudo25. Utilizando esta hipótesis X, Y y Z es posible clasificar tanto las enfermedades como los tratamientos según el componente específico implicado (X, Y o Z).
El epitelio conjuntival, sin embargo, es un epitelio autorrenovable con un gran recambio celular, cuyas células madre van diferenciándose en células epiteliales a lo largo de la vida. Su localización ha sido muy controvertida, pero los estudios clínicos señalan que las células madre conjuntivales se localizan en el fórnix y/o en la conjuntiva bulbar26,27. Las técnicas para determinar la localización de estas células se basa en la determinación de células de ciclo lento mediante el marcaje del DNA mitótico, en la estimulación mitótica, en la capacidad proliferativa de las células aisladas en cultivo y en la definición del origen del movimiento celular. Existen estudios que han analizado la distribución de las células madre epiteliales en la conjuntiva bulbar mediante los movimientos homeostáticos y las mitosis de las células epiteliales en esta zona. Se ha observado que estas células epiteliales de la conjuntiva bulbar eran activas mitóticamente, pero permanecían estacionarias, lo que indicaba su capacidad de autorregeneración así como una distribución uniforme de estas células madre en toda la conjuntiva bulbar28.
Marcadores histológicos de las células madre corneales
El objetivo funcional del epitelio limbocorneal es proporcionar una capa celular intrínsecamente transparente, lisa, altamente hidrofílica, impermeable a la penetración de soluciones fisiológicas y que proteja del ataque de gérmenes patógenos. Estas propiedades dependen de la adquisición de ciertas características así como de la organización celular de una variedad de componentes citoplasmáticos, citoesqueléticos y de membrana que se van adquiriendo conforme se va produciendo la diferenciación celular.
Los métodos para identificar las células madre limbocorneales son indirectos. Todavía no se han establecido marcadores directos de las células madre limbocorneales pero existen evidencias clínicas y experimentales de la localización de estas células madre en la región limbar8,29-33. Arpitha y col34 sugirieron que las células madre limbocorneales probablemente estén constituidas por un grupo de células pequeñas con un cociente núcleo/citoplasma elevado y altos niveles de la proteína celular p63. Se han propuesto una serie de marcadores celulares asociados a las células madre epiteliales de tejidos oculares y no oculares. La proteína p63 parece tener un doble papel en la formación del epitelio, el iniciar la estratificación epitelial durante el desarrollo y el mantener el potencial proliferativo35. Los principales marcadores propuestos se clasifican en tres grupos:
– Proteínas nucleares como el factor de transcripción p63.
– Proteínas de la membrana celular de transmembrana, incluyendo integrinas (b1, a6, etc), receptores (receptor del factor de crecimiento epidérmico o EGFR) y transportadores resistentes a fármacos como el ABCG223,36,37.
– La expresión variable de keratinas en las células epiteliales permite la separación de las poblaciones celulares dentro del epitelio corneal según su nivel de diferenciación. Las citoqueratinas 3 y 12 son marcadores de diferenciación corneal38. La citoqueratina 3 se expresa en el epitelio suprabasal del limbo y en el epitelio corneal completo, pero no se expresa en el epitelio basal del limbo ni en la conjuntiva bulbar adyacente. El epitelio basal limbar tampoco expresa la citoqueratina 12, que sí se expresa en el epitelio limbar suprabasal y en el epitelio corneal completo8,12. La mayoría de células limbares basales están desprovistas de conexinas y proteínas de unión gap. Las uniones gap son un tipo de comunicación específica entre dos células formada por la agrupación local de canales intercelulares de poros grandes (unos 2 nm). Estos canales están constituidos por polipéptidos de tipo conexina, que son identificados según su peso molecular39. La falta de comunicación intercelular entre las células limbares basales refleja la necesidad de un medio ambiente intracelular específico para mantener su indiferenciación8. La membrana basal constituye un componente fundamental en los estímulos exógenos que controlan el crecimiento y la diferenciación de las células limbocorneales. La composición de la membrana se ve directamente afectada por la actividad secretora de las células epiteliales que se apoyan en ella36.
Chen y col23 estudiaron los marcadores de los epitelios corneal y limbar con inmunohistoquímica, PCR (Polymerase Chain Reaction) e hibridación in situ. Observaron cómo el p63, una proteína nuclear, se detectaba en el núcleo de la capa basal del epitelio limbar, pero no en la mayoría de las células epiteliales corneales ni de la capa suprabasal limbar. Este marcador es indicativo de una alta capacidad proliferativa. Kawakita y col no observaron la expresión de p63 en las células basales y en algunas células suprabasales de la córnea normal del conejo. Experimentos in vitro muestran cómo las células madre producen altos niveles de p63 mientras que las células TAC no producen niveles detectables de p63 con técnicas de Western blot. También se ha demostrado cómo el ABCG2, proteína transportadora, se encuentra en la membrana celular y en el citoplasma de la mayoría de las células basales del epitelio limbar, expresándose primariamente el gen ABCG2 en las células basales limbares36,41,42. Igualmente se ha comprobado una alta expresión de integrina a9. También se encontraron otros marcadores, aunque no de forma exclusiva, en esta capa basal del epitelio limbar, como con los marcadores anteriores. Estos son la integrina b1, el EGFR y la citoqueratina 19, que se expresan también en todas las capas del epitelio corneal. Por último, también se ha observado cómo ciertos marcadores no se expresaban o se expresaban mínimamente en el epitelio basal limbar; son la conexina 43, las citoqueratinas 3 y 12, la nestina, la caderina E y la involucrina. Algunos de estos marcadores son específicos del epitelio corneal diferenciado. Se han encontrado en algunos estudios células que coexpresan dos componentes de filamentos celulares intermedios, la vimentina y la keratina 19. Sin embargo, todavía no está claro si se trata de células madre limbocorneales o de células TAC8.
Otros autores, como Watanabe y col36 investigaron si la capa basal del epitelio limbar contiene una subpoblación celular presente en muchos tipos de células madre específicas y que se conoce como células con fenotipo SP (Side Population). Estas células se identifican por su capacidad para captar la tinción Hoechst 33342 y expresan el marcador ABCG236,43-47. El porcentaje medio de células SP en el epitelio limbar a partir de 6 estudios independientes es de 0,29%. Otros estudios, como el de Unemoto y col45, encontraron un 0,4% de células SP en la población epitelial limbar. Los autores comprobaron que la expresión del ABCG2 es significativamente mayor en la subpoblación de células con fenotipo SP que en las que no presentan ese fenotipo. El ABCG2 se expresa en las células epiteliales limbares basales, pero no en las células epiteliales corneales. Este estudio concluye que el marcador ABCG2 puede servir como un marcador de la superficie de células madre del epitelio corneal aunque su función permanece aún desconocida. Actualmente, el ABCG2 parece ser el marcador de superficie celular más útil en la identificación y aislamiento de las células madre limbocorneales48. También se ha demostrado la existencia de esta población de células SP en el limbo de conejo, caracterizada por ser una población indiferenciada y por aumentar su tasa de replicación en casos de cicatrización corneal central49.
Las células madre limbocorneales, originadas embrionariamente en el ectodermo neural, parecen tener propiedades neurológicas in vitro, lo que ha sido demostrado por la reacción inmunohistoquímica a receptores de neurotransmisores (GABA, dopamina, serotonina, etc)50,51.
Harkin y col52 han estudiado la expresión de citoqueratinas y del marcador p63 en cultivos autólogos de células epiteliales limbares. Encontraron una marcada expresión de las citoqueratinas 14 y 19 en las células limbares basales asociada a muy bajos niveles de expresión de citoqueratina 3 y a una alta expresión de p63. Los bajos niveles de citoqueratina 3 confirman la baja diferenciación celular de este epitelio limbar mientras que existe una coexpresión de las citoqueratinas 14 y 19 y del p63, lo que se asocia a un gran potencial proliferativo de las células epiteliales limbares in vitro52. Previamente, Koizumi y col realizaron cultivos directamente sobre membrana amniótica o disociándolos tras su expansión ex vivo53. Esta expansión ex vivo proporciona un número suficiente de células para realizar varios trasplantes evitando la realización de nuevas biopsias. La disociación se lleva a cabo con tripsina al 0,25% y con EDTA 1mM. Aunque el papel exacto de los fibroblastos no está claro, la supervivencia y proliferación de las células progenitoras epiteliales se ve potenciada por interacciones con las células derivadas del ratón y con los productos secretados por dichas células.
Insuficiencia límbica: diagnóstico y caracterización
La insuficiencia límbica es la pérdida parcial o total de las células madre limbocorneales. La etiología de la insuficiencia límbica (IL) puede relacionarse con un microambiente estromal insuficiente para mantener las funciones de las células madre corneales o, en la mayoría de los casos, con factores externos que destruyen las células madre limbares como agresiones químicas o térmicas, radiaciones ultravioletas o ionizantes, síndrome de Stevens-Johnson, penfigoide ocular cicatricial, cirugías, crioterapias, utilización de lentes de contacto o infecciones microbianas54. Incluso, se ha descrito el desarrollo de una insuficiencia límbica completa como complicación del tratamiento con mitomicina C tópica55. Otras veces, lo que ocurre es una hipofunción de estas células, con una pérdida gradual de la función de las células limbocorneales debida a un soporte estromal insuficiente. Esto ocurre en casos de aniridia, queratitis asociada a trastornos endocrinos, queratopatía neurotrófica, inflamaciones limbares crónicas o causas idiopáticas56.
La destrucción parcial o completa del epitelio limbar lleva a una cicatrización anormal del epitelio corneal, dando lugar a una serie de signos clínicos. Los principales signos son la invasión del epitelio conjuntival o conjuntivalización corneal, la vascularización y la inflamación crónica del estroma corneal54,57. Otros signos son un reflejo corneal irregular, una pérdida de las empalizadas de Vogt y una córnea de grosor variable58. La conjuntivalización corneal supone una pérdida del epitelio limbar como barrera entre los epitelios corneal y conjuntival. En condiciones normales, la invasión corneal por parte del epitelio conjuntival vecino es impedida por las células limbares. La conjuntivalización corneal produce una tinción anómala con fluoresceína. Se trata de una tinción tardía debido a que el epitelio conjuntival deja pasar la fluoresceína a diferencia del epitelio corneal. Las células limbares se dañan y el epitelio conjuntival migra sobre el estroma corneal produciendo la conjuntivalización, que se acompaña de vascularización corneal8 (Fig. 5).
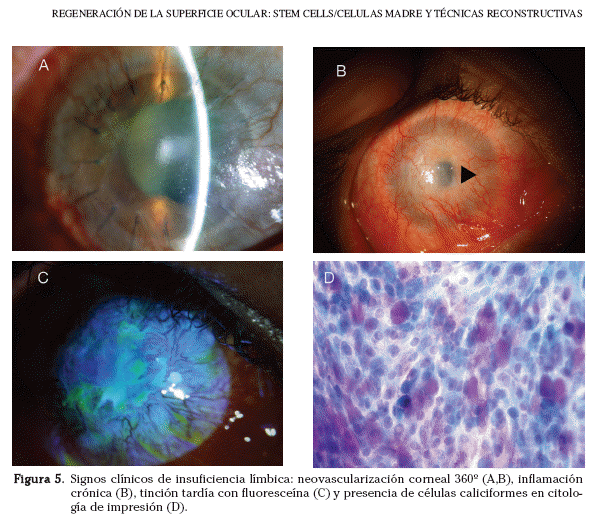
Estas dos manifestaciones clínicas constituyen la definición de insuficiencia límbica. La asociación entre ambos signos clínicos podría estar relacionada con la producción, por parte del epitelio conjuntival, de factores que estimulan la inflamación estromal y la vascularización corneal. El tercer signo clínico que completa la triada de la insuficiencia límbica es una cicatrización enlentecida asociada a erosiones corneales recurrentes. Entre las posibles complicaciones se encuentran la cicatrización, calcificación, ulceración, meelting y perforación corneal.
Los síntomas clínicos son una disminución de la agudeza visual, fotofobia, lagrimeo, blefaroespasmo y episodios recurrentes de dolor junto con inflamación y enrojecimiento crónicos. Los signos típicos de la insuficiencia límbica aguda son fundamentalmente la desaparición de las empalizadas de Vogt59 y el defecto epitelial corneal total que sobrepasa en 2 ó 3 mm el límite limbocorneal. En la insuficiencia límbica crónica, sin embargo, a la desaparición completa de las empalizadas de Vogt se le añaden la neovascularización subepitelial desde el limbo, la queratitis puntata superficial y la hiperpermeabilidad del epitelio que recubre la córnea53.
Hay casos en los que, a pesar de aparecer signos clínicos indicativos de IL, no existe una evidencia citológica de IL en la citología de impresión. Se han postulado diversas causas para explicar esta situación: puede que el daño de las células limbocorneales no sea lo suficientemente severo para destruir toda la población celular, que la insuficiencia límbica sea subclínica y empeore posteriormente, que la citología de impresión no sea lo suficientemente sensible o que algunos pacientes no manifiesten la IL56.
La insuficiencia límbica puede presentarse de forma localizada (parcial) o difusa (total). El diagnóstico de la insuficiencia límbica es muy importante y es determinante del tratamiento a seguir59. En estos pacientes, la queratoplastia lamelar y la queratoplastia completa proporcionan sólo el reemplazo temporal del epitelio corneal del receptor pero no reconstruyen la función limbar. Es necesario llevar a cabo el trasplante autólogo o alogénico de limbo para conseguir la reconstrucción de la superficie corneal. Este tratamiento puede combinarse con una queratoplastia simultánea o posterior.
Insuficiencia límbica parcial
En estos casos la insuficiencia límbica afecta sólo a parte del limbo. El diagnóstico también se basa en la pérdida de las empalizadas de Vogt, y, en algunos casos, en la presencia de células caliciformes en la superficie corneal, observadas en la citología de impresión. Se ha demostrado como el trasplante de membrana amniótica ayuda a la regeneración de la superficie corneal en casos de insuficiencia límbica parcial sin necesidad de llevar a cabo un trasplante de células madre limbocorneales60. Esto apoya la teoría de que la membrana amniótica ayuda a preservar y expandir la población de células madre limbocorneales remanente en casos de insuficiencia límbica parcial60,61. Otros autores también han utilizado la traslocación limbar ipsilateral en casos de insuficiencia límbica parcial secundaria a causticación por álcalis. Esta técnica consiste en trasplantar una zona sana de tejido limbar superior a la zona causticada del mismo ojo, sin tener que recurrir al ojo contralateral62.
Insuficiencia límbica completa
En la insuficiencia límbica completa (ILC), la afectación ocurre en todo el limbo corneal. La detección clínica de la ILC es importante porque estos pacientes son malos candidatos para el trasplante de córnea convencional. La citología de impresión es una técnica sencilla, no invasiva, que se ha demostrado útil en el diagnóstico y monitorización de la ILC. En estos pacientes, se ha propuesto como alternativa a la queratoplastia convencional el trasplante de células madre autólogas asociado al injerto de membrana amniótica. Es necesario el trasplante de una fuente autóloga o alogénica de las células madre limbocorneales que se han perdido63. La membrana amniótica parece preservar las células madre limbocorneales y retener sus propiedades in vivo, actuando como soporte ideal en el cultivo de estas células64. El objetivo de este tratamiento es promover la reepitelización corneal, conseguir un epitelio estable, prevenir la neovascularización y restaurar la transparencia corneal. El injerto de membrana amniótica asociado al alotrasplante de limbo ayuda a la regeneración de un microambiente estromal perilímbico adecuado para soportar el tejido limbar trasplantado. Según Tseng y col los resultados son igual de satisfactorios independientemente de que el trasplante de membrana amniótica se haga antes o en un mismo tiempo que el trasplante de limbo60. El trasplante alogénico presenta muchos problemas asociados, principalmente el alto riesgo de rechazo del injerto debido al gran número de antígenos HLA-DR y células de langerhans en el injerto. De ahí que sean necesarias altas dosis de inmunosupresión durante largos periodos de tiempo8.
Tratamiento quirúrgico en la insuficiencia limbica
Queratoplastia
La queratoplastia penetrante es una técnica quirúrgica que consiste en la extirpación de un botón corneal en el receptor y su sustitución por un botón corneal de un donante, que se sutura a la córnea residual del receptor con una sutura suelta (Fig. 6) o continua. La queratoplastia es indispensable para la reconstrucción del daño estromal severo, pero sólo estimula la reepitelización corneal completa si existen células madre limbocorneales epiteliales en el ojo dañado. Las células limbares pueden ser estimuladas para migrar y proliferar por los efectos paracrinos de las células del donante, como ocurre con los cultivos de queratinocitos de la piel. En ausencia de células epiteliales limbocorneales, la queratoplastia produce reepitelización corneal a partir de células conjuntivales, es decir, una conjuntivalización corneal. En estos pacientes, el trasplante autólogo de células limbares previo es la única posibilidad de cicatrización corneal normal. Este procedimiento implica el trasplante de injertos limbares del ojo sano al ojo dañado.

Trasplante de membrana amniótica
La membrana amniótica, es decir, el amnion, es la capa más interna de la membrana placentaria y está constituida por una membrana basal gruesa y un estroma avascular65. El trasplante de membrana amniótica proporciona un buen sustrato que favorece la cicatricación del epitelio a la superficie corneal. La membrana basal de la membrana amniótica estimula la migración y adhesión de las células epiteliales. La presencia de su matriz estromal avascular disminuye la inflamación, la neovascularización y la fibrosis66. La membrana amniótica produce una mejoría clínica de la fotofobia y el dolor, facilita la epitelización más rápida, ayudando a restaurar una superficie epitelial corneal normal65. Constituye también un sustrato ideal para restaurar el nicho estromal de las células limbocorneales epiteliales así como para conseguir la expansión de estas células67. Ésto se ve favorecido por su carencia de componentes vasculares y por su gruesa membrana basal constituída por colágeno IV y V58.
Trasplante de limbo
La regeneración de esta población limbocorneal epitelial es necesaria para la recuperación de la visión y de la superficie corneal normal cuando existe un déficit de las células madre limbocorneales. Kenyon y Tseng68, en 1989, demostraron la utilidad del trasplante de limbo en casos de insuficiencia límbica completa secundaria a causticaciones térmicas o químicas. Estos hallazgos confirmaron la utilidad del epitelio limbar como fuente de células madre en la regeneración de esta población celular destruida69. Mediante este procedimiento se trasplantan células madre viables procedentes del propio individuo (autotrasplante límbico) o de un donante (alotrasplante límbico) en casos de afectación bilateral. La técnica descrita por ellos consistía en el trasplante de dos fragmentos limbocorneales grandes, de 6-7 mm de arco de limbo, extirpados del ojo sano. El trasplante de limbo no debe hacerse en la fase aguda tras la causticación debido a la alta posibilidad de fracaso asociada a la gran inflamación, debe esperarse varios meses para llevar a cabo el tratamiento70. El autotrasplante de limbo es el primer tratamiento quirúrgico en causticación química o térmica de la córnea unilateral, tras el cual puede llevarse a cabo la queratoplastia en los casos de opacidad estromal71.
Trasplante de células limbares
La pérdida limbocorneal que implica el trasplante de limbo supone un riesgo de desarrollar insuficiencia límbica para el ojo sano. Una forma de minimizar el daño del ojo donante es la expansión in vitro de células epiteliales limbares para su posterior implante. Las células madre limbocorneales expandidas ex vivo pueden reconstruir con éxito la superficie corneal en casos de insuficiencia límbica completa unilatera72. Existen diversas técnicas para llevar a cabo el trasplante de células limbares. El objetivo de este tratamiento es extirpar el tejido corneal anómalo y el pannus corneal del receptor y trasplantar una nueva fuente de células epiteliales corneales. Si el trasplante es eficaz la córnea del receptor queda recubierta por un epitelio corneal sano generado a partir de las células trasplantadas. Se ha demostrado cómo una biopsia limbar de 1 mm2 es suficiente para obtener las células necesarias para regenerar un epitelio corneal sano y cubrir la superficie limbocorneal. Esto significaría una mínima pérdida de células madre limbocorneales en el ojo sano16. El tejido del donante puede proceder del ojo contralateral (autotrasplante), cuando éste está sano, o de un donante (alotrasplante), si la afectación es bilateral. Se necesita un soporte tisular para trasplantar las células ya que éstas no pueden trasplantarse de forma aislada. Están descritos diferentes soportes para el cultivo celular, el más frecuente es la membrana amniótica73-75, pero también se han usado otros como el colágeno76 o el gel de fibrina77. Diversos estudios experimentales han demostrado que durante la expansión ex vivo sobre membrana amniótica se preservan la tasa de replicación, la cinética del ciclo celular y las características fenotípicas de las células progenitoras del epitelio limbar y conjuntival78. La membrana amniótica estimula la migración del epitelio corneal sobre ella promoviendo una fuerte adhesión entre las células basales corneales y el estroma amniótico53. Parece ser que esta posible interacción entre las células limbocorneales y la matriz de la membrana amniótica podría tener un papel fundamental en el mantenimiento de las características de las células madre limbocorneales. Las células expandidas se adhieren fuertemente a la membrana amniótica formándose unas estructuras entre el epitelio y la membrana amniótica similares a la membrana basal78.
Las células madre del epitelio limbocorneal expandidas y preservadas en el cultivo sobre membrana amniótica dan lugar a un epitelio estratificado de fenotipo indiferenciado similar al epitelio limbar in vivo79.
Recientemente, Hiti y col80 han descrito como tratamiento de la insuficiencia límbica completa la realización de una queratoplastia lamelar-penetrante combinada con el trasplante de células limbocorneales y la han llevado a cabo en 6 pacientes. Esta queratoplastia lamelar-penetrante consiste en una queratoplastia lamelar periférica asociada a una queratoplastia penetrante central. Vajpayee y col81 ya habían descrito previamente la utilidad de la queratoplastia lamelar de gran diámetro en el tratamiento de causticaciones químicas, consiguiéndose una superficie ocular estable debido al aporte de células madre limbocorneales.
Bibliografía
1. Mimura T, Yamagami S, Yokoo S, Araie M, Amano S. Comparison of rabbit corneal endothelial cell precursors in the central and peripheral cornea. Invest Ophthalmol Vis Sci 2005; 46: 3645-3648. [ Links ]
2. Krenzer KL, Freddo TF. Cytokeratin expression in normal human bulbar conjunctiva obtained by impression cytology. Invest Ophthalmol Vis Sci 1997; 38: 142-152. [ Links ]
3. Wirtschafter JD, Ketcham JM, Weinstock RJ, Tabesh T, McLoon LK. Mucocutaneous junction as the major source of replacement palpebral conjunctival epithelial cells. Invest Ophthalmol Vis Sci 1999; 40: 3138- 31346. [ Links ]
4. Wei ZG, Wu RL, Lavker RM, Sun TT. In vitro growth and differentiation of rabbit bulbar, fornix, and palpebral conjunctival epithelia. Implications on conjunctival epithelial transdifferentiation and stem cells. Invest Ophthalmol Vis Sci 1993; 34: 1814-1828. [ Links ]
5. Wei ZG, Lin T, Sun TT, Lavker RM. Clonal analysis of the in vivo differentiation potential of keratinocytes. Invest Ophthalmol Vis Sci 1997; 38: 753-761. [ Links ]
6. Meller D, Dabul V, Tseng SC. Expansion of conjunctival epithelial progenitor cells on amniotic membrane. Exp Eye Res 2002; 74: 537-545. [ Links ]
7. Gipson IK. The epithelial basement membrane zone of the limbus. Eye 1989; 3: 132-140. [ Links ]
8. Daniels JT, Dart JK, Tuft SJ, Khaw PT. Corneal stem cells in review. Wound Repair Regen 2001; 9: 483-494. [ Links ]
9. Schermer A, Galvin S, Sun TT. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J Cell Biol 1986; 103: 49-62. [ Links ]
10. Lavker RM, Tseng SC, Sun TT. Corneal epithelial stem cells at the limbus: looking at some old problems from a new angle. Exp Eye Res 2004; 78: 433-446. [ Links ]
11. Lehrer MS, Sun TT, Lavker RM. Strategies of epithelial repair: modulation of stem cell and transit amplifying cell proliferation. J Cell Sci 1998; 111: 2867-2875. [ Links ]
12. Chen JJ, Tseng SC. Corneal epithelial wound healing in partial limbal deficiency. Invest Ophthalmol Vis Sci 1990; 31: 1301-1314. [ Links ]
13. Dua HS, Azuara-Blanco A. Limbal stem cells of the corneal epithelium. Surv Ophthalmol 2000; 44: 415-425. [ Links ]
14. Huang AJ, Tseng SC. Corneal epithelial wound healing in the absence of limbal epithelium. Invest Ophthalmol Vis Sci 1991; 32: 96-105. [ Links ]
15. Sun TT, Lavker RM. Corneal epithelial stem cells: past, present, and future. J Investig Dermatol Symp Proc 2004; 9: 202-207. [ Links ]
16. Pellegrini G, Traverso CE, Franzi AT, Zingirian M, Cancedda R, De Luca M. Long-term restoration of damaged corneal surfaces with autologous cultivated corneal epithelium. Lancet 1997; 349: 990-993. [ Links ]
17. Kinoshita S, Adachi W, Sotozono C et al. Characteristics of the human ocular surface epithelium. Prog Retin Eye Res 2001; 20: 639-673. [ Links ]
18. Boulton M, Albon J. Stem cells in the eye. Int J Biochem Cell Biol 2004; 36: 643-657. [ Links ]
19. Suda T, Arai F, Shimmura S. Regulation of stem cells in the niche. Cornea 2005; 24: S12-17. [ Links ]
20. Potten CS, Loeffler M. Stem cells: attributes, cycles, spirals, pitfalls and uncertainties. Lessons for and from the crypt. Development 1990; 110: 1001-1020. [ Links ]
21. Ahmad I. Stem cells: new opportunities to treat eye diseases. Invest Ophthalmol Vis Sci 2001; 42: 2743-2748. [ Links ]
22. Adhikary G, Crish JF, Bone F, Gopalakrishnan R, Lass J, Eckert RL. An involucrin promoter AP1 transcription factor binding site is required for expression of involucrin in the corneal epithelium in vivo. Invest Ophthalmol Vis Sci 2005; 46: 1219-1227. [ Links ]
23. Chen Z, de Paiva CS, Luo L, Kretzer FL, Pflugfelder SC, Li DQ. Characterization of putative stem cell phenotype in human limbal epithelia. Stem Cells 2004; 22: 355-366. [ Links ]
24. Thoft RA, Friend J. The X, Y, Z hypothesis of corneal epithelial maintenance. Invest Ophthalmol Vis Sci 1983; 24: 1442-1443. [ Links ]
25. Davanger M, Evensen A. Role of the pericorneal papillary structure in renewal of corneal epithelium. Nature 1971; 229: 560-561. [ Links ]
26. Lavker RM, Wei ZG, Sun TT. Phorbol ester preferentially stimulates mouse fornical conjunctival and limbal epithelial cells to proliferate in vivo. Invest Ophthalmol Vis Sci 1998; 39: 301-307. [ Links ]
27. Lavker RM, Sun TT. Epithelial stem cells: the eye provides a vision. Eye 2003; 17: 937-942. [ Links ]
28. Nagasaki T, Zhao J. Uniform distribution of epithelial stem cells in the bulbar conjunctiva. Invest Ophthalmol Vis Sci 2005; 46: 126-132. [ Links ]
29. Charukamnoetkanok P. Corneal stem cells: bridging the knowledge gap. Semin Ophthalmol 2006; 21: 1-7. [ Links ]
30. Chee KY, Kicic A, Wiffen SJ. Limbal stem cells: the search for a marker. Clin Experiment Ophthalmol 2006; 34: 64-73. [ Links ]
31. Lindberg K, Brown ME, Chaves HV, Kenyon KR, Rheinwald JG. In vitro propagation of human ocular surface epithelial cells for transplantation. Invest Ophthalmol Vis Sci 1993; 34: 2672-2679. [ Links ]
32. Romano AC, Espana EM, Yoo SH, Budak MT, Wolosin JM, Tseng SC. Different cell sizes in human limbal and central corneal basal epithelia measured by confocal microscopy and flow cytometry. Invest Ophthalmol Vis Sci 2003; 44: 5125-5129. [ Links ]
33. Hirao A, Arai F, Suda T. Regulation of cell cycle in hematopoietic stem cells by the niche. Cell Cycle 2004; 3: 1481-1483. Epub 2004 Dec 4. [ Links ]
34. Arpitha P, Prajna NV, Srinivasan M, Muthukkaruppan V. High expression of p63 combined with a large N/C ratio defines a subset of human limbal epithelial cells: implications on epithelial stem cells. Invest Ophthalmol Vis Sci 2005; 46: 3631-3636. [ Links ]
35. Koster MI, Kim S, Mills AA, DeMayo FJ, Roop DR. p63 is the mole cular switch for initiation of an epithelial stratification program. Genes Dev 2004; 18: 126-131. Epub 2004 Jan 16. [ Links ]
36. Watanabe K, Nishida K, Yamato M, Umemoto T, Sumide T, Yamamoto K et al. Human limbal epithelium contains side population cells expressing the ATP-binding cassette transporter ABCG2. FEBS Lett 2004; 565: 6-10. [ Links ]
37. Pajoohesh-Ganji A, Pal-Ghosh S, Simmens SJ, Stepp MA. Integrins in slow-cycling corneal epithelial cells at the limbus in the mouse. Stem Cells 2006; 24: 1075-1086. Epub 2005 Nov 10. [ Links ]
38. Liu CY, Zhu G, Converse R, Kao CW, Nakamura H, Tseng SC et al. Characterization and chromosomal localization of the cornea-specific murine keratin gene Krt1.12. J Biol Chem 1994; 269: 24627-24636. [ Links ]
39. Wolosin JM, Xiong X, Schutte M, Stegman Z, Tieng A. Stem cells and differentiation stages in the limbo-corneal epithelium. Prog Retin Eye Res 2000; 19: 223-255. [ Links ]
40. Kawakita T, Espana EM, He H, Li W, Liu CY, Tseng SC. Intrastromal invasion by limbal epithelial cells is mediated by epithelial-mesenchymal transition activated by air exposure. Am J Pathol 2005; 167: 381-393. [ Links ]
41. Budak MT, Alpdogan OS, Zhou M, Lavker RM, Akinci MA, Wolosin JM. Ocular surface epithelia contain ABCG2-dependent side population cells exhibiting features associated with stem cells. J Cell Sci 2005; 118: 1715-1724. [ Links ]
42. de Paiva CS, Chen Z, Corrales RM, Pflugfelder SC, Li DQ. ABCG2 transporter identifies a population of clonogenic human limbal epithelial cells. Stem Cells 2005; 23: 63-73. [ Links ]
43. Goodell MA, Brose K, Paradis G, Conner AS, Mulligan RC. Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo. J Exp Med 1996; 183: 1797-806. [ Links ]
44. Kim M, Turnquist H, Jackson J, Sgagias H, Yan Y, Gong M et al. The multidrug resistance transporter ABCG2 (breast cancer resistance protein 1) effluxes Hoechst 33342 and is overexpressed in hematopoietic stem cells. Clin Cancer Res 2002; 8: 22-28. [ Links ]
45. Umemoto T, Yamato M, Nishida K, Yang J, Tano Y, Okano T. Limbal epithelial side-population cells have stem cell-like properties, including quiescent state. Stem Cells 2006; 24: 86-94. Epub 2005 Sep 8. [ Links ]
46. Wolosin JM, Budak MT, Akinci MA. Ocular surface epithelial and stem cell development. Int J Dev Biol 2004; 48: 981-991. [ Links ]
47. Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J, Morris JJ et al. The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nat Med 2001; 7: 1028-1034. [ Links ]
48. Schlotzer-Schrehardt U, Kruse FE. Identification and characterization of limbal stem cells. Exp Eye Res 2005; 81: 247-264. [ Links ]
49. Park KS, Lim CH, Min BM, Chung HY, Joo CK, Park CW et al. The side population cells in the rabbit limbus sensitively increased in response to the central cornea wounding. Invest Ophthalmol Vis Sci 2006; 47: 892-900. [ Links ]
50. Seigel GM, Sun W, Salvi R, Campbell LM, Sullivan S, Reidy JJ. Human corneal stem cells display functional neuronal properties. Mol Vis 2003; 9: 159-163. [ Links ]
51. Zhao X, Das AV, Thoreson WB, James J, Wattnem TE, Rodríguez-Sierra J et al. Adult corneal limbal epithelium: a model for studying neural potential of non-neural stem cells/progenitors. Dev Biol 2002; 250: 317-331. [ Links ]
52. Harkin DG, Barnard Z, Gillies P, Ainscough SL, Apel AJ. Analysis of p63 and cytokeratin expression in a cultivated limbal autograft used in the treatment of limbal stem cell deficiency. Br J Ophthalmol 2004; 88: 1154-1158. [ Links ]
53. Koizumi N, Fullwood NJ, Bairaktaris G, Inatomi T, Kinoshita S, Quantock AJ. Cultivation of corneal epithelial cells on intact and denuded human amniotic membrane. Invest Ophthalmol Vis Sci 2000; 41: 2506-2513. [ Links ]
54. Dua HS, Saini JS, Azuara-Blanco A, Gupta P. Limbal stem cell deficiency: concept, aetiology, clinical presentation, diagnosis and management. Indian J Ophthalmol 2000; 48: 83-92. [ Links ]
55. Dudney BW, Malecha MA. Limbal stem cell deficiency following topical mitomycin C treatment of conjunctival-corneal intraepithelial neoplasia. Am J Ophthalmol 2004; 137: 950-951. [ Links ]
56.Puangsricharern V, Tseng SC. Cytologic evidence of corneal diseases with limbal stem cell deficiency. Ophthalmology 1995; 102: 1476-1485. [ Links ]
57. Kruse FE, Reinhard T. Die limbustransplantation zur rekonstruktion der augenoberfläche. Ophthalmologe 2001; 98: 818-831. [ Links ]
58. Gomes JA, Dos Santos MS, Cunha MC, Mascaro VL, Barros Jde N, de Sousa LB. Amniotic membrane transplantation for partial and total limbal stem cell deficiency secondary to chemical burn. Ophthalmology 2003; 110: 466-473. [ Links ]
59. Espana EM, Grueterich M, Romano AC, Touhami A, Tseng SC. Idiopathic limbal stem cell deficiency. Ophthalmology 2002; 109: 2004-2010. [ Links ]
60. Tseng SC, Prabhasawat P, Barton K, Gray T, Meller D. Amniotic membrane transplantation with or without limbal allografts for corneal surface reconstruction in patients with limbal stem cell deficiency. Arch Ophthalmol 1998; 116: 431-441. [ Links ]
61. Anderson DF, Ellies P, Pires RT, Tseng SC. Amniotic membrane transplantation for partial limbal stem cell deficiency. Br J Ophthalmol 2001; 85: 567-575. [ Links ]
62. Nishiwaki-Dantas MC, Dantas PE, Reggi JR. Ipsilateral limbal translocation for treatment of partial limbal deficiency secondary to ocular alkali burn. Br J Ophthalmol 2001; 85: 1031-1033. [ Links ]
63. Levin LA, Ritch R, Richards JE, Borras T. Stem cell therapy for ocular disorders. Arch Ophthalmol 2004; 122: 621-627. [ Links ]
64. Meller D, Pires RT, Tseng SC. Ex vivo preservation and expansion of human limbal epithelial stem cells on amniotic membrane cultures. Br J Ophthalmol 2002; 86: 463-471. [ Links ]
65. Meller D, Pires RT, Mack RJ, Figueiredo F, Heiligenhaus A, Park WC et al. Amniotic membrane transplantation for acute chemical or thermal burns. Ophthalmology 2000; 107: 980-989; discussion 990. [ Links ]
66.Dua HS, Azuara-Blanco A. Amniotic membrane transplantation. Br J Ophthalmol 1999; 83: 748-752. [ Links ]
67. Grueterich M, Espana EM, Touhami A, Ti SE, Tseng SC. Phenotypic study of a case with successful transplantation of ex vivo expanded human limbal epithelium for unilateral total limbal stem cell deficiency. Ophthalmology 2002; 109: 1547-1552. [ Links ]
68. Kenyon KR, Tseng SC. Limbal autograft transplantation for ocular surface disorders. Ophthalmology 1989; 96: 709-722; discussion 722-723. [ Links ]
69. Tsubota K, Satake Y, Kaido M, Shinozaki N, Shimmura S, Bissen-Miyajima H et al. Treatment of severe ocular-surface disorders with corneal epithelial stem-cell transplantation. N Engl J Med 1999; 340: 1697-1703. [ Links ]
70. Basti S, Rao SK. Current status of limbal conjunctival autograft. Curr Opin Ophthalmol 2000; 11: 224-232. [ Links ]
71. Shimazaki J, Shimmura S, Tsubota K. Donor source affects the outcome of ocular surface reconstruction in chemical or thermal burns of the cornea. Ophthalmology 2004; 111: 38-44. [ Links ]
72. Ti SE, Anderson D, Touhami A, Kim C, Tseng SC. Factors affecting outcome following transplantation of ex vivo expanded limbal epithelium on amniotic membrane for total limbal deficiency in rabbits. Invest Ophthalmol Vis Sci 2002; 43: 2584-2592. [ Links ]
73. Tsai RJ, Li LM, Chen JK. Reconstruction of damaged corneas by transplantation of autologous limbal epithelial cells. N Engl J Med 2000; 343: 86-93. [ Links ]
74. Schwab IR, Reyes M, Isseroff RR. Successful transplantation of bioengineered tissue replacements in patients with ocular surface disease. Cornea 2000; 19: 421-426. [ Links ]
75. Koizumi N, Inatomi T, Suzuki T, Sotozono C, Kinoshita S. Cultivated corneal epithelial stem cell transplantation in ocular surface disorders. Ophthalmology 2001; 108: 1569-1574. [ Links ]
76. He YG, Alizadeh H, Kinoshita K, McCulley JP. Experimental transplantation of cultured human limbal and amniotic epithelial cells onto the corneal surface. Cornea 1999; 18: 570-579. [ Links ]
77. Rama P, Bonini S, Lambiase A, Golisano O, Paterna P, De Luca M et al. Autologous fibrin-cultured limbal stem cells permanently restore the corneal surface of patients with total limbal stem cell deficiency. Transplantation 2001; 72: 1478-1485. [ Links ]
78. Grueterich M, Espana E, Tseng SC. Connexin 43 expression and proliferation of human limbal epithelium on intact and denuded amniotic membrane. Invest Ophthalmol Vis Sci 2002; 43: 63-71. [ Links ]
79. Tseng SC, Meller D, Anderson DF, Touhami A, Pires RT, Grüterich M et al. Ex vivo preservation and expansion of human limbal epithelial stem cells on amniotic membrane for treating corneal diseases with total limbal stem cell deficiency. Adv Exp Med Biol 2002; 506: 1323-1334. [ Links ]
80. Hiti B, Tost F, Clemens S. Optische lamellär-perforierende keratoplatik mit stammzelltransplantation bei risikofällen. Ophthalmologie 2006; 103: 523-528. [ Links ]
81. Vajpayee RB, Thomas S, Sharma N, Dada T, Tabin GC. Large-diameter lamellar keratoplasty in severe ocular alkali burns: A technique of stem cell transplantation. Ophthalmology 2000; 107: 1765-1768. [ Links ]
![]() Dirección para correspondencia:
Dirección para correspondencia:
Ana Fernández
Departamento de Oftalmología
línica Universitaria
Avda. Pío XII, 36
31008 Pamplona
E-mail: anafern@gmail.com
Recepción el 12 de julio de 2007
Aceptación provisional 7 de noviembre de 2007
Aceptación definitiva 17 de enero de 2008

