










 Custom services
Custom services
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
La calcificación vascular (CV) es una alteración prevalente en el envejecimiento, que se correlaciona con un aumento en la rigidez de los vasos e incremento del riesgo de muerte cardiovascular 1. En la población general, la progresión y la tasa de CV se han asociado con un aumento de riesgo de fracturas y osteoporosis 2, precediendo las alteraciones vasculares a las alteraciones propias del hueso que se observan más tarde 3. Distintos estudios epidemiológicos han mostrado la relación existente entre las alteraciones del metabolismo óseo, la CV y el aumento de la mortalidad 4.
El mecanismo por el cual se produce la CV resulta complejo. Inicialmente, se pensaba que era un proceso pasivo consistente en una simple precipitación de calcio y fósforo en un microambiente apropiado 5. Sin embargo, en la actualidad se sabe que, además de este proceso pasivo, hay mecanismos activos que discurren a la par que los primeros. Durante estos procesos activos, las células de músculo liso vascular (CMLV), debido a ciertos promotores de la calcificación, sufren un grado mayor de apoptosis, forman vesículas y finalmente cambian su fenotipo de células musculares lisas a células de tipo osteoblasto, induciendo formación de matriz y también atrayendo factores locales determinantes del proceso de mineralización 1.
La vitamina D, que ejerce su principal efecto fisiológico sobre la homeostasis del calcio y del fósforo, ha sido tradicionalmente usada para el tratamiento y prevención del raquitismo y de la osteomalacia 6. Se hidroxila en el riñón por acción de la 25-hidroxivitamina D-1α hidroxilasa dando lugar al calcitriol, que es el metabolito fisiológico más activo de esta hormona 7. Desde hace años se conoce que in vitro el calcitriol a dosis elevadas induce un incremento de la calcificación en CMLV 8, de la actividad de la fosfatasa alcalina y una regulación a la baja del péptido relacionado con la hormona paratiroidea (PTHrP) 9. Estudios posteriores han confirmado el efecto promotor de la calcificación del calcitriol al aumentar el depósito de calcio en la aorta de ratas con función renal normal 10 y al incrementar los niveles de ARN mensajero (ARNm) de las proteínas óseas: Factor de transcripción 2 relacionado con Runt (Runx2), osteocalcina, osteoprotegerina (OPG), receptor activador para el factor nuclear κβ (RANKL) y proteína morfogenética ósea 4 (BMP4) en la aorta de ratas nefrectomizadas 11,12.
Por tanto, el objetivo de este estudio fue evaluar la expresión diferencial de proteínas que se produce con el efecto calcificante de dosis suprafisiológicas de calcitriol en un modelo in vitro de CMLV.
Material y métodos
Inducción de calcificación vascular por calcitriol
La línea celular de músculo liso vascular de aorta de rata, SMAC-R (primary culture DPK-SMAC-R; Pharmakine), fue cultivada a 37ºC en una atmósfera húmeda con 5% de CO2 en Dulbecco’s Modified Eagle Medium (DMEM, Lonza) con 10% de suero fetal bovino (FBS, HyClone® Thermo Scientific), penicilina 100 UI/mL, estreptomicina 100 µg/mL y glutamina 2 mM (Biochrom AG).
Alcanzada la confluencia necesaria (60%, aproximadamente), las células se cultivaron con medio de cultivo DMEM F-12 suplementado con 0,1% de albúmina sérica bovina (BSA) en ausencia o presencia de calcitriol (10-7 M, Sigma-Aldrich) durante diez días, cambiando el medio de cultivo por medio fresco cada 48 horas. Todos los experimentos se llevaron a cabo por triplicado y cada condición en cada experimento se hizo por triplicado.
Determinación de la actividad fosfatasa alcalina
Las CMLV cultivadas en placas de 24 pocillos con o sin calcitriol 10-7 M durante 10 días fueron recogidas, y se cuantificó la actividad de la fosfatasa alcalina midiendo la cantidad de para-nitrofenol fosfato hidrolizado siguiendo las instrucciones del kit utilizado para su determinación (BioAssay Systems).
Inmunocitoquímica
Para evaluar la pérdida del fenotipo vascular, se realizó la inmunocitoquímica de α-actina. Para ello, las CMLV crecieron en un soporte plástico específico para ello (Thermanox) y se expusieron posteriormente a calcitriol 10-7 M durante 10 días. Para ello, se utilizó el anticuerpo monoclonal de ratón frente α-actina (CP-47, Calbiochem) y el kit Dako Real™ EnVision™ Detection System, Peroxidase/DAB+, Rabbit/Mouse (Dako).
Análisis de la expresión génica
Para el análisis de la expresión génica mediante PCR cuantitativa a tiempo real (qPCR) se extrajo previamente el ARN de las células utilizando Tri Reagent (Ambion) siguiendo un protocolo estándar. A partir de 2 µg de ARN total, se obtuvo el ADNc utilizando el kit High Capacity cDNA Reverse Transcription (Applied Biosystems) siguiendo las instrucciones del fabricante. Para cuantificar la expresión relativa de los genes, se utilizó un termociclador modelo Stratagene Mx3005P QPCR System (Agilent Technologies) y reactivos Taqman® (Taqman® Universal PCR Master Mix, Applied Biosystems). Se utilizaron los ensayos de Applied Biosystems correspondientes a los siguientes genes: α-actina (Rn01759928_g1), elastina (Rn01499782_m1), osteocalcina (Rn01455285_g1) y Runx2 (Rn01512296_m1). Los resultados se normalizaron frente al control endógeno 18S (Eukaryotic 18S rRNA endogenous control reagent, Applied Biosystems). La interpretación de los datos se realizó mediante el método de la comparación del ciclo umbral (ΔΔCt) 13.
Estudio proteico
Elecrotroforesis bidimensional de proteínas:
Para el estudio de la expresión diferencial de proteínas por electroforesis bidimensional se cultivaron CMLV durante 10 días en ausencia o presencia de calcitriol 10-7 M. La extracción de las proteínas totales se realizó homogeneizando las células en un tampón de lisis compuesto por urea 7 M, tiourea 2 M y CHAPS 2%. Las proteínas fueron purificadas y desaladas empleando el kit Ready Prep Clean-up (Bio-Rad) y cuantificadas mediante el ensayo de Bradford 14.
El isoelectroenfoque (separación de las proteínas según su punto isoeléctrico) o primera dimensión, se realizó por triplicado con 150 µg de proteínas de cada condición en tiras de poliacrilamida deshidratada de 24 cm con un rango de pH 3-10 no lineal (IPG-Strips, GE Healthcare). 24 horas antes del isoelectroenfoque, las tiras se rehidrataron con DeStreak Rehydration Solution y anfolitos al 0,5% (ambos de GE Healthcare). El isoelectroenfoque fue realizado en un Ettan IPGphor3 (GE Healthcare), y posteriormente las tiras se lavaron con tampón de equilibrado (urea 6 M, Tris-HCL 75 mM, glicerol 30% y SDS 2%) con dos lavados de 15 minutos, el primero con dithiothreitol (DTT) 1% y el segundo con iodoacetamida 2,5%, para la reducción-alquilación de las proteínas.
La segunda dimensión se realizó mediante SDS-PAGE en geles de poliacrilamida al 12% en una cubeta múltiple (GE Healthcare). Una vez terminada la electroforesis, los geles fueron teñidos con nitrato de plata 0,1% y digitalizados con un densitómetro calibrado GS-800, analizando la diferente intensidad de los spots mediante el programa informático PDQuest, ambos de Bio-Rad.
Análisis de la expresión diferencial de proteínas mediante espectrometría de masas:
Los spots expresados de manera diferencial en las dos condiciones de cultivo (ausencia o presencia de calcitriol) fueron recortados del gel, troceados y desteñidos en incubación durante 10 minutos con ferrocianuro potásico 30 mM y tiosulfato sódico 100 mM, y deshidratados en acetonitrilo. Finalmente, fueron digeridos con tripsina 13 ng/µL en bicarbonato amónico 10 mM/ acetonitrilo 10% durante 24 horas a 37ºC. Los péptidos obtenidos de la digestión con tripsina se identificaron mediante cromatografía de líquidos (nanoHPLC, Applied Biosystems) y espectrometría de masas por trampa iónica (Q-TRAP, Applied Biosystems) (LC-MS/MS). Los espectros de masas de las proteínas expresadas diferencialmente fueron identificados usando la base de datos UniProtKB/Swissprot (www.uniprot.org) y el motor de búsqueda Mascot.
Análisis proteico mediante Western blot:
Para la confirmación de los resultados obtenidos en la electroforesis bidimensional se utilizó el análisis Western blot. Para ello se extrajeron las proteínas totales de los cultivos en tampón de lisis RIPA y se cuantificaron mediante el método estándar de Bradford (Bio-Rad).
Los diferentes extractos proteicos (30 µg) se sometieron a electroforesis en geles de poliacrilamida en condiciones desnaturalizantes (SDS-PAGE) 15. Las proteínas se transfirieron a una membrana de PVDF (Difluoruro de polivinilideno) (Hybond™ P, GE Healthcare) que se incubó con el anticuerpo primario correspondiente en la dilución adecuada: frente a OPG (sc-8468, Santa Cruz Biotechnology, dilución 1:1.000); frente a α-actina (CP-47, Calbiochem, dilución 1:1.000) y frente a GAPDH (Gliceraldehído-3-fosfato deshidrogenasa) (sc25778, Santa Cruz Biotechnology, 1:5.000). Los anticuerpos secundarios ligados a peroxidasa fueron los específicos para cada anticuerpo primario (sc-2023, Santa Cruz Biotechnology; y 401215, Calbiochem). Por último, se llevó a cabo la detección mediante el kit ECL Western Blotting Detection (Amersham Biosciences). El revelado se llevó a cabo usando el equipo Chemidoc XRS+ (Bio-Rad). La determinación de la intensidad de las bandas obtenidas en el Western blot se realizó con el programa informático Image Lab (Bio-Rad).
Análisis estadístico
El análisis estadístico se realizó utilizando el software SPSS para Windows 17.0 (SPSS Inc) y se consideraron diferencias significativas aquellas con un p valor menor de 0,05. Los resultados se expresaron como media ± desviación estándar. Las diferencias en la actividad de la fosfatasa alcalina y expresión génica y proteica entre las distintas condiciones de cultivo se evaluaron mediante la t de Student después de comprobar la normalidad de las variables. En el caso de la identificación de proteínas con el programa de búsqueda Mascot, la asunción por el mismo de normalidad de las variables ha impedido evaluar la normalidad o no de las proteínas identificadas.
Resultados
Se analizaron en las CMLV los cambios a nivel fenotípico hacia célula tipo osteoblasto tras mantenerlas en cultivo en presencia o ausencia de calcitriol 10-7 M durante 10 días. Se observó un incremento significativo de la actividad de la fosfatasa alcalina de dos veces en las células expuestas a calcitriol (Figura 1A). A su vez, se observó mediante inmunocitoquímica un descenso en α-actina en las células expuestas a calcitriol, lo que confirma la pérdida del fenotipo muscular con el calcitriol a dosis elevadas (Figura 1B).

Figura 1 A) Actividad de la fosfatasa alcalina (ALP), medida como nmoles de p-nitrofenol fosfato/μg proteína hidrolizados por minuto, en CMLV expuestas a calcitriol durante 10 días. B) Inmunocitoquímica de α-actina en CMLV en ausencia (control) o presencia de calcitriol 10-7 M (CTR 10-7 M) durante 10 días. El círculo rojo se representa a mayor aumento a la derecha para ver la diferencia de tinción. *p<0,05 respecto al control
El estudio por qPCR de genes musculares y óseos mostró que la exposición a una dosis alta de calcitriol disminuyó significativamente un 35% la expresión génica de α-actina, mientras que para la elastina la supresión fue casi total (99%) (Figuras 2A y 2B). En el caso de los genes típicamente óseos, se observó que la exposición a calcitriol incrementó de manera significativa (13 veces) la expresión génica de osteocalcina, siendo este aumento menor, aunque también significativo, en el caso del Runx2 (2,5 veces) (Figuras 2C y 2D).
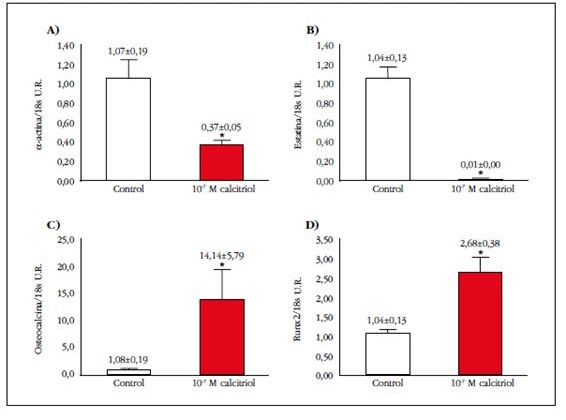
Figura 2 Niveles relativos de ARNm de A) α-actina, B) elastina, C) osteocalcina y D) Runx2 en CMLV en ausencia (control) o presencia de calcitriol 10-7 M (CTR 10-7 M) durante 10 días. *p<0,05 frente a control
Con el fin de comparar el espectro de proteínas expresadas diferencialmente por la exposición de CMLV a calcitriol 10-7 M durante 10 días (grupo CTR 10-7 M) con respecto a CMLV en ausencia de calcitriol (grupo Control), se llevó a cabo un análisis proteómico. Se obtuvieron 334 spots localizados en cada uno de los 6 geles teñidos con nitrato de plata (3 Control y 3 CTR 10-7 M), de entre los cuales 22 presentaron diferencias significativas de expresión (p<0,05). De estos, se identificaron por el buscador Mascot 10 spots que se muestran en la figura 3.

Figura 3 Geles bidimensionales de extractos proteicos de CMLV cultivadas en ausencia (control) o presencia de calcitriol 10-7 M (CTR 10-7 M) durante 10 días. Se señalan aquellas proteínas expresadas diferencialmente respecto a las células control: en rojo las que muestran aumento y en verde las que descienden
De las 10 proteínas que fueron identificadas (Figura 3 y Tabla 1), cabe destacar el aumento de expresión en 6 de ellas, entre las que destaca la superóxido dismutasa mitocondrial, marcador de estrés oxidativo, que experimentó un mayor incremento. El resto de proteínas que aumentó su expresión fueron proteínas del citoesqueleto o relacionadas con él (proteína ácida fibrilar gliar y treonina/serina quinasa tipo Ste20), proteínas que participan en la formación de vesículas (dinaminas), proteínas de membrana (ceramida glucosiltransferasa) y proteínas del inflamasoma (pirinas). Por el contrario, se identificaron 4 proteínas que tras la exposición a calcitriol disminuyeron su expresión, destacando por su mayor descenso la actina citoplasmática 2. El otro marcador muscular cuya expresión se vio también disminuida fue la α-actina de músculo liso aórtico. Las otras dos proteínas cuyas expresiones se vieron disminuidas fueron la prolil-4-hidroxilasa, que participa en la maduración de las fibras de colágeno, y la dipeptidil peptidasa 10 inactiva (DPP10), que forma parte de canales de potasio.
Para confirmar los resultados obtenidos por proteómica se analizó por Western blot la expresión proteica de α-actina, proteína muscular, y de OPG de estirpe ósea. El Western blot mostró que la exposición al calcitriol indujo un descenso en la expresión proteica de α-actina y un incremento de OPG (Figura 4).
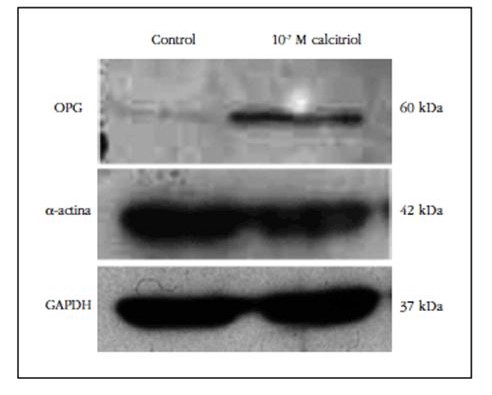
Figura 4 Sobreexpresión de osteoprotegerina (OPG) y descenso de α-actina por Western blot en CMLV expuestas a calcitriol 10-7 M durante 10 días
Tabla 1 Listado de proteínas identificadas por el buscador Mascot expresadas diferencialmente en CMLV cultivadas en ausencia (control) o presencia de calcitriol 10-7 M (CTR 10-7 M) durante 10 días

Se muestran los números de acceso de Swissprot (Nº acc.) y el número de péptidos identificados mediante espectrometría de masas (Nº péptidos). Se representa la media de intensidad de los spots ± desviación estándar de los 3 geles control y de los 3 geles CTR 10-7 M y la tasa de cambio entre ellas (CTR/Control). P calculada mediante t de Student para variables independientes.
Discusión
La investigación en el área de la CV ha despertado un interés creciente debido a su asociación con morbilidad y mortalidad cardiovascular 16. El efecto de la vitamina D como promotor de la CV merece una particular atención, ya que dosis altas de calcitriol se han asociado experimentalmente con un incremento de la CV 9,17. Por ello, se ha analizado in vitro el perfil de expresión diferencial de proteínas de CMLV sometidas a una dosis suprafisiológica de calcitriol, encontrando por vez primera proteínas que no se habían identificado en este proceso de calcificación mediado por calcitriol.
En el presente estudio y de acuerdo con trabajos previos 9,18, se observó un incremento de la actividad de la fosfatasa alcalina y un descenso de la proteína α-actina en las células expuestas a calcitriol. Este efecto puede ser explicado por el aumento de la expresión del factor de transcripción Runx2, el cual ejerce un papel determinante en el cambio fenotípico asociado a la CV 19. También se ha observado un aumento en la transcripción del gen óseo osteocalcina en respuesta al calcitriol y un aumento en la expresión proteica de OPG 20, ambas típicamente óseas. De hecho, la administración de dosis elevadas de calcitriol, similares a las utilizadas en nuestro estudio 9,21, se ha asociado con CV tanto en ratas nefrectomizadas como en ratas con función renal normal 10,21. Por el contrario, dosis bajas de calcitriol y de sus análogos no inducen CV, e incluso pueden tener un efecto protector sobre su desarrollo 22,23.
Paralelamente al aumento en la expresión de factores óseos, se ha observado no solo el descenso en los niveles de proteína α-actina, sino además una disminución en la expresión génica tanto de α-actina como de elastina, ambos marcadores musculares. Dado que las CMLV presentan una elevada plasticidad 24, los promotores de la CV pueden inducir una disminución en la expresión de genes musculares y producir una diferenciación hacia fenotipo óseo, acompañado de un aumento de factores que promueven la calcificación 25.
Tras el análisis por espectrometría de masas de los extractos proteicos obtenidos, se identificaron 10 proteínas. Según donde ejercen su función, las proteínas identificadas se clasificaron en: proteínas del citoesqueleto, que participan en la formación de vesículas, proteínas de membrana, proteínas de matriz extracelular, proteínas del inflamasoma y proteínas relacionadas con el estrés oxidativo. Las proteínas identificadas se distribuyeron mayoritariamente en el citoplasma, pero también en otros orgánulos intracelulares como el retículo endoplasmático (RE), aparato de Golgi y mitocondria.
De las proteínas expresadas diferencialmente e identificadas por LC-MS/MS, seis vieron aumentada su expresión tras el tratamiento con calcitriol respecto al control. La que presentó mayor sobreexpresión fue la superóxido dismutasa o SOD mitocondrial, que es una óxido-reductasa y una de las enzimas antioxidantes más importantes. Dicho aumento podría representar un mecanismo compensador para contrarrestar el daño inducido por el aumento de especies reactivas de oxígeno en el proceso de calcificación 26,27,28.
Otra proteína que se observa aumentada en las CMLV expuestas a calcitriol es la proteína ácida fibrilar gliar, una de las proteínas fibrosas que forman los filamentos intermedios del citoesqueleto intracelular 29. Esta proteína se encuentra en ciertas células estrechamente relacionada con filamentos de vimentina, desmina y periferina, los cuales están implicados en la estructura y función del citoesqueleto. No hay datos que relacionen a la proteína ácida fibrilar gliar con la CV, pero su aumento podría también representar un mecanismo compensador para evitar la desorganización del citoesqueleto que ocurre en el proceso de CV.
En el presente estudio también se observó un aumento en dinamina, que es una GTPasa responsable de la endocitosis en las células eucariotas. Las dinaminas están involucradas principalmente en la escisión de las vesículas recién formadas de la membrana de un compartimento celular. Este hecho las podría implicar en un fenómeno común e inductor de la CV como es la aparición de vesículas matriz, que se forman a partir de células donde se origina mineralización o son el resultado del proceso de apoptosis celular (cuerpos apoptóticos) 30. De hecho, Kashiwakura y cols. han observado que la dinamina es capaz de regular al menos parcialmente la apoptosis inducida por las lipoproteínas de baja densidad oxidadas regulando su endocitosis 31. Un reciente artículo las ha implicado como un mediador del estrés oxidativo en cardiomiocitos, ayudando a frenar la producción de especies reactivas de oxígeno y de apoptosis 32.
Otro grupo de proteínas que aumentó su expresión en CMLV por su exposición a calcitriol fue la treonina/serina quinasa tipo Ste20. Se trata de enzimas implicadas en la orientación y organización de los microtúbulos del huso durante la mitosis 33. Se conoce que esta proteína es un regulador transcripcional de la quinasa poloide Plk1 en el músculo liso 34. Un reciente estudio ha observado un incremento de la expresión de esta proteína en aortas de individuos de edad avanzada, sugiriendo que promueve la inestabilidad de los microtúbulos y filamentos de actina 35.
La glucosilceramida transferasa (GlcT-1) también aumentó su expresión proteica. Esta proteína se integra en el metabolismo de los esfingolípidos, interviniendo dentro de este en la transferencia de una molécula de glucosa para la glicosilación de la ceramida (acilesfingosina), dando lugar a los compuestos glicolipídicos denominados cerebrósidos y que son importantes componentes de la membrana celular del tejido muscular. Si bien no hay datos en la literatura que relacionen estas enzimas con el proceso de calcificación vascular, existe un trabajo clásico que encuentra la acumulación de glicoesfingolípidos en pacientes fallecidos con placa aterosclerótica, lo que podría sugerir un mecanismo patogénico de alteración del tejido vascular 36.
Cabe destacar el aumento de una proteína denominada pirina que forma parte del inflamasoma. El inflamasoma es un conjunto de multiproteínas citosólicas que permite la activación de las caspasas proinflamatorias, las cuales transforman el precursor de la interleukina-1β (pro-IL-1 β) a la forma activa (IL-1 β), lo que conduce a una poderosa respuesta inflamatoria 37. Recientemente, Wen y cols. han descrito que, para que se produzca la calcificación de las CMLV, se requiere que el inflamasoma se active 38.
Asimismo, en concordancia con trabajos previos publicados llevados a cabo en modelos in vitro de disfunción endotelial y proteómica 39, nuestros resultados mostraron un descenso en la expresión de proteínas estructurales y una desregulación de las proteínas del citoesqueleto en las CMLV expuestas a calcitriol a dosis suprafisiológicas. La actina del músculo liso aórtico y la actina citoplasmática 2 son proteínas que juegan un papel fundamental en la arquitectura y motilidad celular. Este hecho podría interpretarse como una pérdida del fenotipo muscular de las CMVL como consecuencia de la exposición a altas concentraciones de calcitriol.
La CV es un proceso patológico regulado que se asemeja a la osteogénesis. Cuando las CMLV de la media se exponen a un estímulo calcificante mantienen su capacidad para diferenciarse en células tipo osteoblasto o condrocito, expresando distintas proteínas óseas, produciendo vesículas de matriz y componentes de la matriz extracelular con propensión a la mineralización 40.
En nuestros experimentos, además de componentes de la matriz extracelular, se detectó un descenso en proteínas del retículo endoplasmático rugoso (RER) implicadas en la maduración del colágeno. La prolil-4-hidroxilasa es una enzima que participa en la hidroxilación post-traslacional de la prolina del procolágeno y cuyo descenso inhibe la formación de la triple hélice. Si bien no hay datos de esta enzima sobre la CV, la ausencia de prolil-3-hidroxilasa con la que comparte el mecanismo de hidroxilación de la prolina en el colágeno da lugar a una hipermineralización de la matriz ósea 41.
Por ultimo, una proteína que ha mostrado un descenso es la dipeptidil peptidasa 10 inactiva (DPP10). Esta proteína, que es un importante componente neuronal de los canales de potasio, puede actuar como una chaperona interactuando con otras importantes moléculas de señalización, como hsp90 y proteínas asociadas, pudiendo modular la apoptosis 42. Por lo tanto, el descenso observado en nuestro estudio en las células expuestas a calcitriol podría ser una consecuencia del cambio en el fenotipo celular observado.
Los resultados de este estudio experimental in vitro tienen la limitación de no ser aplicables directamente a humanos, pero sí que deberían alertar del efecto que dosis elevadas de calcitriol, empleadas como tratamiento del hiperparatiroidismo secundario en la enfermedad renal crónica, podrían tener sobre el proceso de calcificación vascular. Aunque en la población normal el riñón tendría capacidad para eliminar los excesos de calcitriol, en su mayoría derivados del tratamiento con 25-hidroxivitamina D y de su conversión renal y extra-renal a calcitriol, cuando el deterioro de la función renal se agrava por el envejecimiento se debería alertar a los clínicos de seguir un control muy estricto, para evitar efectos nocivos que podría tener un calcitriol elevado, aumentando la hipercalcemia e hiperfosforemia y su posterior depósito en el tejido vascular.
La identificación de los spots de proteínas está limitada por la cantidad y la calidad (poca concentración de sales, ácidos nucleicos, lípidos, etc.) del extracto proteínico obtenido, lo cual depende directamente de la eficacia del método de extracción 43. En nuestro trabajo, además de las proteínas descritas anteriormente identificadas con una alta fiabilidad, según la puntuación obtenida en las diferentes bases de datos de identificación de proteínas, se identificaron también otras proteínas con una baja puntuación que podría ser explicada atendiendo a diferentes razones 44. Por un lado, la concentración de algunas moléculas puede ser muy inferior respecto a otras mayoritarias en los spots cortados. Por otro lado, algunas proteínas pueden haber experimentado modificaciones post-traduccionales o proteólisis, pudiendo alterar la disponibilidad de los péptidos para su identificación. Por último, hay que tener en cuenta que la caracterización del perfil proteico de las células se ha llevado a cabo en un modelo de calcificación que se prolongó hasta los 10 días. Es posible que, en modelos donde la calcificación se establezca en periodos de hasta 3 semanas, los niveles de algunas proteínas pudieran ser más altos y observarse mayores diferencias.
Por tanto, y a modo de resumen, podemos afirmar que hemos objetivado una disminución del fenotipo vascular y un aumento del óseo en CMLV sometidas a una dosis suprafisiológica de calcitriol. Estos cambios fenotípicos dan lugar a un perfil diferencial de proteínas, con aumento de la expresión de proteínas que participan en la depuración de radicales libres y que forman parte del citoesqueleto como un posible mecanismo compensador al proceso de calcificación. También se observó un aumento de proteínas que participan en la formación de vesículas de matriz, así como en un aumento en la respuesta inflamatoria, ambos procesos inherentes a la calcificación vascular. La pérdida de fenotipo muscular se vio representada por descensos en la expresión de proteínas típicamente musculares. Estos resultados deberían ser ratificados en modelos animales con vistas a su utilidad clínica en la prevención de calcificaciones vasculares en población general.

