










My SciELO
 Custom services
Custom servicesServices on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de Osteoporosis y Metabolismo Mineral
On-line version ISSN 2173-2345Print version ISSN 1889-836X
Rev Osteoporos Metab Miner vol.9 n.4 Madrid Nov./Dec. 2017
https://dx.doi.org/10.4321/s1889-836x2017000400005
Originals
Effect of supra-physiological calcitriol doses on protein expression of vascular smooth muscle cells
1 Servicio de Metabolismo Óseo y Mineral - Instituto Reina Sofía de Investigación Nefrológica - Red de Investigación Renal del Instituto de Salud Carlos III (REDinREN del ISCIII) - Universidad de Oviedo - Oviedo (España)
2 Laboratorio de Bioquímica - Hospital 12 de Octubre - Madrid (España)
3 Laboratorio de Medicina - Hospital Universitario Central de Asturias - Oviedo (España)
Introduction
Vascular calcification (VC) is a prevalent alteration in aging, which has been linked to an increase in vessel stiffness and an increased risk of cardiovascular death 1. In the general population, progression and VC rate have been associated with an increased risk of fractures and osteoporosis 2, preceding vascular alterations to bone alterations observed later 3. Different epidemiological studies have shown the relationship between alterations in bone metabolism, VC and increased mortality 4.
The mechanism by which VC occurs is complex. Initially, it was thought to be a passive process consisting of a simple precipitation of calcium and phosphorus in an appropriate microenvironment 5. However, it is now known that, in addition to this passive process, there are active mechanisms that run along with the former. During these active processes, vascular smooth muscle cells (VSMC), due to certain calcification promoters, suffer a greater degree of apoptosis, form vesicles and finally change their phenotype of smooth muscle cells to osteoblast-like cells, inducing matrix formation and also attracting local factors that determine the mineralization process 1.
Vitamin D, which exerts its main physiological effect on calcium and phosphorus homeostasis, has traditionally been used to treat and prevent rickets and osteomalacia 6. It is hydroxylated in the kidney by the action of 25-hydroxyvitamin D-1α hydroxylase giving rise to calcitriol, which is the most active physiological metabolite of this hormone 7. For years, high-dose calcitriol has been known to induce increased calcification in VSMC 8, alkaline phosphatase activity and reduced regulation of parathyroid hormone-related peptide (PTHrP) 9. Further studies have confirmed the calcification promoting effect of calcitriol by increasing calcium deposition in the aorta of the rat model with normal renal function 10 and by increasing Messenger RNA levels (mRNA) of bone proteins: Runt-related transcription factor 2 (Runx2), osteocalcin, osteoprotegerin (OPG), activator receptor for nuclear factor κβ (RANKL) and bone morphogenetic protein 4 (BMP4) in the aorta of nephrectomized rats 11,12.
Therefore, this study aimed to evaluate the differential expression of proteins produced with the calcifying effect of supra-physiological doses of calcitriol in an in vitro model of VSMC.
Material and methods
Induction of calcitriol vascular calcification
The rat aorta vascular smooth muscle cell line, SMAC-R (primary culture DPK-SMAC-R; Pharmakine), was cultured at 37°C in a humid atmosphere with 5% CO2 in Dulbecco's Modified Eagle Medium (DMEM, Lonza) with 10% fetal bovine serum (FBS, HyClone® Thermo Scientific), 100 IU/mL penicillin, 100 µg/mL streptomycin and 2 mM glutamine (Biochrom AG).
Upon reaching the necessary confluence (approximately 60%), the cells were cultured with DMEM F-12 culture medium supplemented with 0.1% bovine serum albumin (BSA) in the absence or presence of calcitriol (10-7 M, Sigma- Aldrich) for ten days, changing the culture with fresh medium every 48 hours. All experiments were carried out in triplicate and each condition in each experiment was done in triplicate.
Determination of alkaline phosphatase activity
VSMCs cultured in 24-well plates with or without 10-7 M calcitriol for 10 days were collected, and alkaline phosphatase activity was quantified by measuring the amount of hydrolyzed para-nitrophenol phosphate following the instructions of the kit used for its determination (BioAssay Systems).
Immunocytochemistry
To evaluate the vascular phenotype loss, the immunocytochemistry of α-actin was carried out. In order to do so, the VSMCs were grown on a specific plastic support (Thermanox) and then exposed to 10-7 M calcitriol for 10 days. The mouse monoclonal antibody against α-actin (CP-47, Calbiochem) and the Dako Real™ EnVision™ Detection System, Peroxidase/DAB+, Rabbit/Mouse (Dako) were used for this purpose.
Analysis of gene expression
For the analysis of gene expression by quantitative real-time PCR (qPCR) RNA was previously extracted from the cells using Tri Reagent (Ambion) following a standard protocol. From 2 µg of total RNA, the cDNA was obtained using the High Capacity cDNA Reverse Transcription kit (Applied Biosystems) following the manufacturer's instructions. To quantify the relative genetic expression, a thermocycler model Stratagene Mx3005P QPCR System (Agilent Technologies) and Taqman® reagents (Taqman® Universal PCR Master Mix, Applied Biosystems) were used. We used the Applied Biosystems assays corresponding to the following genes: α-actin (Rn01759928_g1), elastin (Rn01499782_m1), osteocalcin (Rn01455285_g1) and Runx2 (Rn01512296_m1). The results were normalized against 18S endogenous control (Eukaryotic 18S rRNA endogenous control reagent, Applied Biosystems). The interpretation of the data was performed using the threshold cycle comparison method (ΔΔCt) 13.
Protein study
Two-dimensional protein electrotrophrosis:
For the study of differential protein expression by two-dimensional electrophoresis, VSMC was cultured for 10 days in the absence or presence of 10-7 M calcitriol. Total protein extraction was performed by homogenizing the cells in a lysis buffer composed of 7 M urea, 2 M thiourea and 2% CHAPS. Proteins were purified and desalted using the Ready Prep Clean-up kit (Bio-Rad) and quantified by the Bradford assay 14.
The isoelectric focusing (separation of the proteins according to their isoelectric point) or first dimension was carried out in triplicate with 150 µg of proteins of each condition in strips of 24 cm dehydrated polyacrylamide with a non-linear pH range 3-10 (IPG-Strips, GE Healthcare). At 24 hours prior to isoelectric focusing, the strips were rehydrated with DeStreak Rehydration Solution and 0.5% ampholytes (both from GE Healthcare). The isoelectric focusing was performed on Ettan IPGphor3 (GE Healthcare), and the strips were then washed with equilibration buffer (6 M urea, 75 mM Tris-HCL, 30% glycerol and 2% SDS) with two 15-minute washes, First with dithiothreitol (DTT) 1% and the second with 2.5% iodoacetamide, for reduction-alkylation of proteins.
The second dimension was performed by SDS-PAGE on 12% polyacrylamide gels in a multiple cuvette (GE Healthcare). After the electrophoresis, the gels were stained with 0.1% silver nitrate and digitized with a calibrated densitometer GS-800, analyzing the different intensity of the spots using the PDQuest software, both of Bio-Rad.
Analysis of differential protein expression by mass spectrometry:
Differentially expressed spots in the two culture conditions (absence or presence of calcitriol) were cut from the gel, sliced and washed out in incubation for 10 minutes with 30 mM potassium ferrocyanide and 100 mM sodium thiosulfate, and dehydrated in acetonitrile. Finally, they were digested with 13 ng/µL trypsin in 10 mM ammonium bicarbonate/10% acetonitrile for 24 hours at 37°C. Peptides obtained from trypsin digestion were identified by liquid chromatography (nanoHPLC, Applied Biosystems) and ion trap mass spectrometry (Q-TRAP, Applied Biosystems) (LC-MS/MS). Mass spectra of differentially expressed proteins were identified using the UniProtKB/Swissprot database (www.uniprot.org) and the Mascot search engine.
Protein analysis by Western blot:
Western blot analysis was used to confirm the results obtained in two-dimensional electrophoresis. To this end, the total proteins were extracted from the cultures in RIPA lysis buffer and quantified by the Bradford standard method (Bio-Rad).
The different protein extracts (30 µg) were electrophoresed on polyacrylamide gels under denaturing conditions (SDS-PAGE) 15. The proteins were transferred to a PVDF (Hybond™ P, GE Healthcare) membrane which was incubated with the corresponding primary antibody at the appropriate dilution: against OPG (sc-8468, Santa Cruz Biotechnology, 1:1,000); against α-actin (CP-47, Calbiochem, 1:1,000 dilution) and against GAPDH (scycealdehyde-3-phosphate dehydrogenase) (sc25778, Santa Cruz Biotechnology, 1:5,000). Peroxidase-linked secondary antibodies were specific for each primary antibody (sc-2023, Santa Cruz Biotechnology; and 401215, Calbiochem). Finally, the detection was carried out by the ECL Western Blotting Detection kit (Amersham Biosciences). The development was carried out using the Chemidoc XRS+ kit (Bio-Rad). The determination of the intensity of the bands obtained in the Western blot was performed with the Image Lab software (Bio-Rad).
Statistic analysis
Statistical analysis was carried out using SPSS software for Windows 17.0 (SPSS Inc) and significant differences were considered with p less than 0.05. Results were expressed as mean ± standard deviation. Differences in alkaline phosphatase activity and gene and protein expression between the different culture conditions were assessed using Student's t test after checking the normality of the variables. In the case of the identification of proteins with the Mascot search program, the assumption of the variables’ normality has hindered assessing the normality or not of the identified proteins.
Results
Changes in the phenotypic level to the osteoblast-like cell were analyzed in the VSMC after being maintained in culture in the presence or absence of 10-7 M calcitriol for 10 days. A significant increase in alkaline phosphatase activity was observed twice in cells exposed to calcitriol (Figure 1A). In addition, a decrease in α-actin in cells exposed to calcitriol was observed by immunocytochemistry, confirming the loss of muscle phenotype with calcitriol at high doses (Figure 1B).
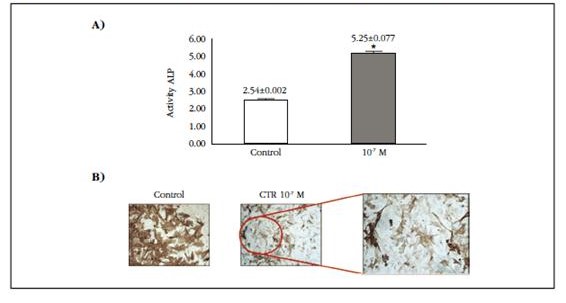
Figure 1 A) Alkaline phosphatase activity (ALP), measured as nmol of p-nitrophenol phosphate/μg protein hydrolyzed per minute, in VCMV exposed to calcitriol for 10 days. B) Immunocytochemistry of α-actin in VSMC in absence (control) or presence of 10-7 M calcitriol (10-7 M CTR) for 10 days. The red circle is depicted at higher magnification on the right to see the difference in staining. * P<0.05 with respect to the control
The qPCR study of muscle and bone genes showed that exposure to a high dose of calcitriol significantly decreased α-actin gene expression by 35%, whereas for elastin suppression was almost total (99%) (Figures 2A and 2B). In the case of typically osseous genes, exposure to calcitriol significantly increased (13 fold) the gene expression of osteocalcin, this increase being smaller, but also significant, in the case of Runx2 (2.5 fold) (Figures 2C and 2D).
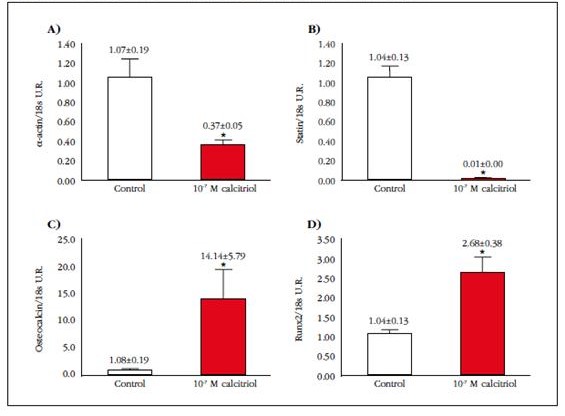
Figure 2 Relative levels of mRNA of A) α-actin, B) elastin, C) osteocalcin and D) Runx2 in VSMC in absence (control) or presence of 10-7 M calcitriol (10-7 M CTR) for 10 days. *P<0.05 vs control
In order to compare the spectrum of differentially expressed proteins by VSMC exposure to 10-7 M calcitriol for 10 days (10-7 M CTR group) with respect to VSMC in the absence of calcitriol (control group), proteomic analysis was carried out. There were 334 spots located on each of the 6 gels stained with silver nitrate (3 Control and 3 CTR 10-7 M), of which 22 presented significant differences in expression (p <0.05). Of these, Mascot 10 spots were identified by the search engine that are shown in figure 3.
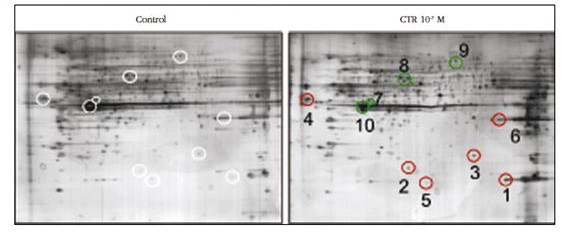
Figure 3 Two-dimensional gels of VSMC protein extracts cultured in absence (control) or presence of 10-7 M calcitriol (10-7 M CTR) for 10 days. Those proteins expressed differentially with respect to the control cells are indicated: in red those that show increase and in green those that descend
Of the 10 proteins that were identified (Figure 3 and Table 1), we should highlight the increase in expression in 6 of them, among which is the mitochondrial superoxide dismutase, a marker of oxidative stress, which experienced a greater increase. The other proteins that increased its expression were cytoskeletal or related proteins (glial fibrillary acid and threonine/serine kinase type Ste20), proteins involved in vessel formation (dynamins), membrane proteins (ceramide glucosyltransferase) and proteins of the inflammasome (pyrins). In contrast, we identified 4 proteins that after their exposure to calcitriol decreased their expression, with the highest decrease in cytoplasmic actin 2. The other muscle marker whose expression was also diminished was α-actin of aortic smooth muscle. The other two proteins whose expressions were diminished were prolyl-4-hydroxylase, involved in the maturation of collagen fibers, and inactive dipeptidyl peptidase 10 (DPP10), which is part of potassium channels.
To confirm the results obtained by proteomics the protein expression of α-actin, muscle protein, and OPG of bone strain was analyzed by Western blot analysis. Western blot analysis showed that exposure to calcitriol induced a decrease in α-actin protein expression and an increase in OPG (Figure 4).

Figure 4 Overexpression of osteoprotegerin (OPG) and decrease of α-actin by Western blot in VSMC exposed to 10-7 M calcitriol for 10 days
Table 1 List of proteins identified by the Mascot searcher differentially expressed in VSMC cultured in absence (control) or presence of 10-7 M calcitriol (10-7 M CTR) for 10 days

The accession numbers of Swissprot (Nº acc.) And the number of peptides identified by mass spectrometry (Nº peptides) are shown. The average intensity of the spots ± standard deviation of the 3 control gels, the 3 gels CTR 10-7 M and the exchange between them (CTR/Control) is represented. P calculated by Student t for independent variables.
Discussion
Research into CV has aroused increasing interest due to its association with cardiovascular morbidity and mortality 16. The effect of vitamin D as a promoter of CV deserves particular attention, since high doses of calcitriol have been associated experimentally with an increase in CV 9,17. Therefore, the differential expression profile of VSMC proteins subjected to a supra-physiological dose of calcitriol has been analyzed in vitro, finding for the first-time proteins that had not been identified in this process of calcification mediated by calcitriol.
In the present study and according to previous studies 9,18, an increase in alkaline phosphatase activity and a decrease in α-actin protein were observed in cells exposed to calcitriol. This effect can be explained by the increased expression of the transcription factor Runx2, which plays a decisive role in the phenotypic change associated with CV 19. An increase in the transcription of the osteocalcin bone gene in response to calcitriol and an increase in the protein expression of OPG, both typically bone, have also been observed 20. In fact, administration of high doses of calcitriol, similar to those used in our study 9,21, has been associated with CV in both nephrectomized rats and in rats with normal renal function 10,21. In contrast, low doses of calcitriol and its analogs do not induce CV, and may even have a protective effect on its development 22,23.
Parallel to the increase in the expression of bone factors, not only the decrease in α-actin protein levels, but also a decrease in the gene expression of both α-actin and elastin, both muscle markers, have been observed. Since VCAMs show high plasticity 24, VC promoters can induce a decrease in the expression of muscle genes and lead to a differentiation into the bone phenotype, accompanied by an increase in factors that promote calcification 25.
After mass spectrometric analysis of the protein extracts obtained, 10 proteins were identified. Depending on where they exert their function, the identified proteins were classified as: cytoskeleton proteins, involved in the formation of vesicles, membrane proteins, extracellular matrix proteins, inflammatory proteins and proteins related to oxidative stress. The identified proteins were mostly distributed in the cytoplasm, but also in other intracellular organelles such as the endoplasmic reticulum (ER), Golgi apparatus and mitochondria.
Of the differentially expressed proteins identified by LC-MS/MS, six showed increased expression after treatment with calcitriol compared to the control. The one with the greatest overexpression was superoxide dismutase or mitochondrial SOD, which is an oxide-reductase and one of the most important antioxidant enzymes. Such increase could represent a compensatory mechanism to counteract the damage induced by the increase of reactive oxygen species in the calcification process 26,27,28.
Another protein that is observed to be increased in the VCMVs exposed to calcitriol is the glial fibrillary acidic protein, one of the fibrous proteins that form the intermediate filaments of the intracellular cytoskeleton 29. This protein is found in certain cells closely related to filaments of vimentin, desmin and periferin, which are involved in the structure and function of the cytoskeleton. There are no data linking the glial fibrillary acidic protein with the CV, but its increase could also represent a compensatory mechanism to avoid the disorganization of the cytoskeleton that occurs in the CV process.
In the present study, we also observed an increase in dynamin, which is a GTPase responsible for endocytosis in eukaryotic cells. Dynamins are mainly involved in the excision of newly formed vesicles from the membrane of a cell compartment. This fact could involve them in a common phenomenon and inducer of CV as is the appearance of matrix vesicles, which are formed from cells where mineralization originates or are the result of the process of cellular apoptosis (apoptotic bodies) 30. In fact, Kashiwakura et al. have observed that dynamin is capable of at least partially regulating apoptosis induced by oxidized low-density lipoproteins by regulating its endocytosis 31. A recent article has implicated them as a mediator of oxidative stress in cardiomyocytes, helping to slow the production of reactive oxygen species and apoptosis 32.
Another group of proteins that increased their expression in VSMC by their exposure to calcitriol was the Ste20 threonine/serine kinase. These enzymes are involved in the orientation and organization of spindle microtubules during mitosis 33. It is known that this protein is a transcriptional regulator of the poloid kinase Plk1 in smooth muscle 34. A recent study has observed an increase in the expression of this protein in aortas of elderly individuals, suggesting that it promotes the instability of microtubules and actin filaments 35.
Glucosylceramide transferase (GlcT-1) also increased its protein expression. This protein is integrated in the metabolism of sphingolipids, intervening in the transfer of a molecule of glucose for the glycosylation of ceramide (acylsphingosine), giving rise to glycolipid compounds called cerebrosides and which are important components of the cell membrane of muscle tissue. Although there are no data in the literature that relate these enzymes to the vascular calcification process, there is a classic work that finds the accumulation of glycosphingolipids in patients who died with atherosclerotic plaque, which could suggest a pathogenic mechanism of vascular tissue alteration 36.
It is possible to emphasize the increase of a protein called pyrin that forms part of the inflammasome. This is a set of cytosolic multi-proteins that allows the activation of proinflammatory caspases which transform the precursor of interleukin-1β (pro-IL-1 β) to the active form (IL-1 β), leading to a powerful inflammatory response 37. Recently, Wen et al. have described that, for the calcification of VSMC, the inflammatory response is required 38.
Likewise, in agreement with previous published works carried out in in vitro models of endothelial dysfunction and proteomics 39, our results showed a decrease in the expression of structural proteins and a deregulation of the cytoskeletal proteins in the VCLCs exposed to calcitriol at supraphysiological doses. Aortic smooth muscle actin and cytoplasmic actin 2 are proteins that play a key role in cell architecture and motility. This could be interpreted as a loss of the VSMC muscle phenotype as a consequence of exposure to high concentrations of calcitriol.
CV is a regulated pathological process that resembles osteogenesis. When the VSMC of the mean are exposed to a calcifying stimulus, they maintain their ability to differentiate into osteoblast or chondrocyte cells, expressing different bone proteins, producing matrix vesicles and components of the extracellular matrix with propensity for mineralization 40.
In our experiments, in addition to components of the extracellular matrix, a decrease in proteins of the rough endoplasmic reticulum (RER) involved in collagen maturation was detected. Prolyl-4-hydroxylase is an enzyme that participates in post-translational hydroxylation of proline proline and whose decrease inhibits the formation of the triple helix. Although there is no data of this enzyme on CV, the absence of prolyl-3-hydroxylase with which it shares the hydroxylation mechanism of proline in the collagen results in hypermineralization of the bone matrix 41.
Finally, a protein that has shown a decrease is inactive dipeptidyl peptidase 10 (DPP10). This protein, which is an important neuronal component of the potassium channels, can act as a chaperone interacting with other important signaling molecules, such as hsp90 and associated proteins, and can modulate apoptosis 42. Therefore, the decrease observed in our study in cells exposed to calcitriol could be a consequence of the change in the observed cellular phenotype.
The results of this in vitro experimental study have the limitations of not being directly applicable to humans, but should warn of the effect that high doses of calcitriol, used as a treatment of secondary hyperparathyroidism in chronic kidney disease, may have on the vascular calcification. Although in the normal population the kidney would be able to eliminate excess calcitriol, mostly derived from treatment with 25-hydroxyvitamin D and its renal and extra-renal conversion to calcitriol, when deterioration of renal function is aggravated by aging should alert clinicians to follow a very strict control, to avoid harmful effects that could have a high calcitriol, increasing hypercalcemia and hyper-phosphoremia and its subsequent deposition in vascular tissue.
Identification of protein spots is limited by the quantity and quality (low concentration of salts, nucleic acids, lipids, etc.) of the protein extract obtained, which depends directly on the efficiency of the extraction method 43. In our work, in addition to the proteins described above identified with a high reliability, according to the score obtained in the different databases of protein identification, other proteins were also identified with a low score that could be explained considering different reasons 44. On the one hand, the concentration of some molecules may be much lower than others in the cut spots. On the other hand, some proteins may have undergone post-translational modifications or proteolysis, which may alter the availability of the peptides for identification. Finally, it should be taken into account that the characterization of the protein profile of the cells has been carried out in a calcification model that lasted up to 10 days. It is possible that, in models where calcification is established in periods of up to 3 weeks, the levels of some proteins could be higher and greater differences observed.
Therefore, and as a summary, we can affirm that we have objectified a decrease in vascular phenotype and an increase of bone in VSMC subjected to a supra-physiological dose of calcitriol. These phenotypic changes give rise to a differential protein profile, with increased expression of proteins involved in free radical scavenging and forming part of the cytoskeleton as a possible compensatory mechanism to the calcification process. It was also observed an increase in proteins involved in the formation of matrix vesicles, as well as in an increase in the inflammatory response, both processes inherent to vascular calcification. The loss of muscle phenotype was represented by declines in the expression of typically muscle proteins. These results should be ratified in animal models with a view to their clinical utility in the prevention of vascular calcifications in the general population.
REFERENCES
1. Giachelli CM. Vascular calcification mechanisms. J Am Soc Nephrol. 2004;15:2959-64. [ Links ]
2. Naves M, Rodriguez-Garcia M, Diaz-Lopez JB, Gomez-Alonso C, Cannata-Andia JB. Progression of vascular calcifications is associated with greater bone loss and increased bone fractures. Osteoporos Int. 2008;19:1161-6. [ Links ]
3. Szulc P, Kiel DP, Delmas PD. Calcifications in the abdominal aorta predict fractures in men: MINOS study. J Bone Miner Res. 2008;23:95-102. [ Links ]
4. Rodriguez Garcia M, Naves Diaz M, Cannata Andia JB. Bone metabolism, vascular calcifications and mortality: associations beyond mere coincidence. J Nephrol. 2005;4:458-63. [ Links ]
5. Schinke T, Karsenty G. Vascular calcification-a passive process in need of inhibitors. Nephrol Dial Transplant. 2000;15:1272-4. [ Links ]
6. Hollick, MF. Resurrection of vitamin D deficiency and rickets. J Clin Invest. 2006;116:2062-72. [ Links ]
7. Cannata-Andia JB, Gomez Alonso C. Vitamin D deficiency: a neglected aspect of disturbed calcium metabolism in renal failure. Nephrol Dial Transplant. 2002;17:1875-8. [ Links ]
8. Inoue T, Kawashima H. 1,25-Dihydroxyvitamin D3 stimulates 45Ca2+-uptake by cultured vascular smooth muscle cells derived from rat aorta. Biochem Biophys Res Commun. 1988;152:1388-94. [ Links ]
9. Jono S, Nishizawa Y, Shioi A, Morii H. 1,25-Dihydroxyvitamin D3 increases in vitro vascular calcification by modulating secretion of endogenous parathyroid hormone-related peptide. Circulation. 1998;98:1302-6. [ Links ]
10. Bas A, Lopez I, Perez J, Rodriguez M, Aguilera-Tejero E. Reversibility of calcitriol-induced medial artery calcification in rats with intact renal function. J Bone Miner Res. 2006;21:484-90. [ Links ]
11. Mizobuchi M, Finch JL, Martin DR, Slatopolsky E. Differential effects of vitamin D receptor activators on vascular calcification in uremic rats. Kidney Int. 2007;72:709-15. [ Links ]
12. Panizo S, Cardus A, Encinas M, Parisi E, Valcheva P, López-Ongil S, et al. RANKL increases vascular smooth muscle cell calcification through a RANK-BMP4-dependent pathway. Circ Res. 2009;104:1041-8. [ Links ]
13. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method Methods. 2001;25:402-8. [ Links ]
14. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248-54. [ Links ]
15. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1990;227:680-5. [ Links ]
16. Blacher J, Guerin AP, Pannier B, Marchais SJ, London GM. Arterial calcifications, arterial stiffness, and cardiovascular risk in end-stage renal disease. Hypertension. 2001;38:938-42. [ Links ]
17. Rebsamen MC, Sun J, Norman AW, Liao JK. 1alpha,25-dihydroxyvitamin D3 induces vascular smooth muscle cell migration via activation of phosphatidylinositol 3-kinase. Circ Res. 2000;91:17-24. [ Links ]
18. Shalhoub V, Shatzen EM, Ward SC, Young JI, Boedigheimer M, Twehues L, et al. Chondro/osteoblastic and cardiovascular gene modulation in human artery smooth muscle cells that calcify in the presence of phosphate and calcitriol or paricalcitol. J Cell Biochem. 2010;111;911-21. [ Links ]
19. Sun Y, Byon CH, Yuan K, Chen J, Mao X, Heath JM, et al. Smooth muscle cell-specific runx2 deficiency inhibits vascular calcification. Circ Res. 2012;111:543-52. [ Links ]
20. Gori F, Hofbauer LC, Dunstan CR, Spelsberg TC, Khosla S, Riggs BL. The expression of osteoprotegerin and RANK ligand and the support of osteoclast formation by stromal-osteoblast lineage cells is developmentally regulated. Endocrinology. 2000;141:4768-76. [ Links ]
21. Wu-Wong JR, Noonan W, Ma J, Dixon D, Nakane M, Bolin AL, et al. Role of phosphorus and vitamin D analogs in the pathogenesis of vascular calcification. J Pharmacol Exp Ther. 2006;318:90-8. [ Links ]
22. Li X, Speer MY, Yang H, Bergen J. Giachelli CM. Vitamin D receptor activators induce an anticalcific paracrine program in macrophages: requirement of osteopontin. Arterioscler Thromb Vasc Biol. 2010;30:321-6. [ Links ]
23. Aoshima Y, Mizobuchi M, Ogata H, Kumata C, Nakazawa A, Kondo F, et al. Vitamin D receptor activators inhibit vascular smooth muscle cell mineralization induced by phosphate and TNF-alpha. Nephrol Dial Transplant. 2012;27:1800-6. [ Links ]
24. Katoh Y, Periasamy M. Growth and differentiation of smooth muscle cells during vascular development. Trends Cardiovasc Med. 1996;6:100-6. [ Links ]
25. Tukaj C, Kubasik-Juraniec J, Kraszpulski M. Morphological changes of aortal smooth muscle cells exposed to calcitriol in culture. Med Sci Monit. 2000;6:668-74. [ Links ]
26. Byon CH, Javed A, Dai Q, Kappes JC, Clemens TL, Darley-Usmar VM, et al. Oxidative stress induces vascular calcification through modulation of the osteogenic transcription factor Runx2 by AKT signaling. J Biol Chem. 2008;283:15319-27. [ Links ]
27. Sutra T, Morena M, Bargnoux AS, Caporiccio B, Canaud B, Cristol JP. Superoxide production: a procalcifying cell signalling event in osteoblastic differentiation of vascular smooth muscle cells exposed to calcification media. Free Radic Res. 2008;42:789-97. [ Links ]
28. Roman-Garcia P, Barrio-Vazquez S, Fernandez-Martin JL, Ruiz-Torres MP, Cannata-Andia, JB. Natural antioxidants and vascular calcification: a possible benefit. J Nephrol. 2011;24:669-72. [ Links ]
29. Lewis GP, Matsumoto B, Fisher SK. Changes in the organization and expression of cytoskeletal proteins during retinal degeneration induced by retinal detachment. Invest Ophthalmol Vis Sci. 1995;36:2404-16. [ Links ]
30. Kapustin AN, Chatrou MLL, Drozdov I, Zheng Y, Davidson SM, Soong D, et al. Vascular smooth muscle cell calcification is mediated by regulated exosome secretion. Circ Res. 2015;116:1312-23. [ Links ]
31. Kashiwakura Y, Watanabe M, Kusumi N, Sumiyoshi K, Nasu Y, Yamada H, et al. Dynamin-2 regulates oxidized low-density lipoprotein-induced apoptosis of vascular smooth muscle cell. Circulation. 2004;110:3329-34. [ Links ]
32. Gao D, Yang J, Wu Y, Wang Q, Wang Q, Lai EY, et al. Targeting dynamin 2 as a novel pathway to inhibit cardiomyocyte apoptosis following oxidative stress. Cell Physiol Biochem. 2016;39:2121-34. [ Links ]
33. Zhapparova ON, Fokin AI, Vorobyeva NE, Bryantseva SA, Nadezhdina ES. Ste20-like protein kinase SLK (LOSK) regulates microtubule organization by targeting dynactin to the centrosome. Mol Biol Cell. 2013;24,3205-14. [ Links ]
34. Li J, Wang R, Gannon OJ, Rezey AC, Jiang S, Gerlach BD, et al. Polo-like kinase 1 regulates vimentin phosphorylation at Ser-56 and contraction in smooth muscle. J Biol Chem. 2016;291:23693-703. [ Links ]
35. Fu Z, Wang M, Everett A, Lakatta E, Van Eyk J. Can proteomics yield insight into aging aorta? Proteomics Clin Appl. 2013;7 477-89. [ Links ]
36. Chatterjee SB, Dey S, Shi WY, Thomas K, Hutchins GM. Accumulation of glycosphingolipids in human atherosclerotic plaque and unaffected aorta tissues. Glycobiology. 1997;7:57-65. [ Links ]
37. Drenth JPH, van der Meer JWM. The Inflammasome - a linebacker of innate defense. N Engl J Med. 2006;355:730-2. [ Links ]
38. Wen C, Yang X, Yan Z, Zhao M, Yue X, Cheng X, et al. Nalp3 inflammasome is activated and required for vascular smooth muscle cell calcification. Int J Cardiol. 2013;168:2242-7. [ Links ]
39. Carbó C, Arderiu G, Escolar G, Fusté B, Cases A, Carrascal M, et al. Differential expression of proteins from cultured endothelial cells exposed to uremic versus normal serum. Am J Kidney Dis. 2008;51:603-12. [ Links ]
40. Neven E, Persy V, Dauwe S, De Schutter T, De Broe ME, D'Haese PC. Chondrocyte rather than osteoblast conversion of vascular cells underlies medial calcification in uremic rats. Arterioscler Thromb Vasc Biol. 2010;30:1741-50. [ Links ]
41. Fratzl-Zelman N, Bächinger HP, Vranka JA, Roschger P, Klaushofer K, Rauch F. Bone matrix hypermineralization in prolyl-3 hydroxylase 1 deficient mice. Bone. 2016;85:15-22. [ Links ]
42. Tsaprouni L, Ito K, Cookson WO, Moffatt MF, Barnes PJ, Adcock IM. Functional genomics of DPP10. Protein characterization and its association with asthma and COPD. Am J Respir Crit Care Med. 2007;175:A387. [ Links ]
43. Gil-Dones F, Martín-Rojas T, López-Almodóvar LF, Juárez-Tosina R, de la Cuesta F, Álvarez-Llamas G, et al. Obtención de un protocolo óptimo para el análisis proteómico de válvulas aórticas humanas sanas y estenóticas. Rev Esp Cardiol. 2010;63:46-53. [ Links ]
44. Bagnato C, Thumar J, Mayya V, Hwang SI, Zebroski H, Claffey KP, et al. Proteomics analysis of human coronary atherosclerotic plaque: a feasibility study of direct tissue proteomics by liquid chromatography and tandem mass spectrometry. Mol Cell Proteomics. 2007;6:1088-102. [ Links ]
Acknowledgments:
This paper has been made possible thanks to the funding obtained by the FEIOMM grant for basic research 2010. This work has also been partially funded with the help of the National R&D&I Plan 2008-2011, State Plan for R&I 2013-2016, Carlos III Health Institute (ISCIII) – European Regional Development Fund (PI13/00014), Science, Technology and Innovation Plan 2013- 2017 of the Principality of Asturias (GRUPIN14-028), Foundation for Development in Asturias Institute for Applied Scientific Research and Technology (FICYT), Reina Sofía Institute for Nephrology Research, Renal Íñigo Álvarez de Toledo Foundation, RETIC RedInRen of ISCIII – European Regional Development Fund (RD06/0016/1013, RD12/0021/1023 and RD16/0009/0017), by the Sociedad Asturiana Fomento Metabólicas Investigaciones.
FundingThis paper has been made possible thanks to the funding obtained by the FEIOMM grant for basic research 2010
FundingScience, Technology and Innovation Plan 2013-2017 of the Principality of Asturias (GRUPIN14-028)
FundingFoundation for Development in Asturias Institute for Applied Scientific Research and Technology (FICYT)
Received: March 31, 2017; Accepted: May 16, 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
