










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista de Osteoporosis y Metabolismo Mineral
versión On-line ISSN 2173-2345versión impresa ISSN 1889-836X
Rev Osteoporos Metab Miner vol.11 no.2 Madrid abr./jun. 2019 Epub 20-Ene-2020
https://dx.doi.org/10.4321/s1889-836x2019000200002
ORIGINALS
The Wnt/β-catenin pathway decreases the amount of osteoclasts in the bone and promotes its apoptosis
1Departamento de Medicina Interna - Hospital Universitario Marqués de Valdecilla - Santander (España)
2Instituto de Biomedicina y Biotecnología de Cantabria (IBBTEC) - Consejo Superior de Investigaciones Científicas - Universidad de Cantabria - Sociedad para el Desarrollo de Cantabria (CSIC-UC-SODERCAN) - Parque Científico y Tecnológico de Cantabria (PCTCAN) - Santander (España)
6Instituto de Biomedicina y Biotecnología de Cantabria (IBBTEC) - Consejo Superior de Investigaciones Científicas - Universidad de Cantabria - Sociedad para el Desarrollo de Cantabria (CSIC-UC-SODERCAN) - Parque Científico y Tecnológico de Cantabria (PCTCAN) - Santander (España)
3Instituto de Investigación Marqués de Valdecilla (IDIVAL) - Santander (España)
4Division de Endocrinología y Metabolismo - Centro para Osteoporosis y Enfermedades Metabólicas Óseas - Universidad de Arkansas de Ciencias Médicas - Sistema de Salud Central para Veteranos de Arkansas - Little Rock - Arkansas (EE.UU.)
5Departamento de Anatomía y Biología Celular - Facultad de Medicina - Universidad de Cantabria - Santander (España)
The activation of Wnt/βcatenin signaling in cells of the osteoblastic lineage leads to an increase in bone mass through a dual mechanism: increasing osteoblastogenesis and decreasing osteoclastogenesis. The predominance of one mechanism over another depends on the maturational state of the osteoblast in which βcatenin accumulation occurs. The activation of Wnt/βcatenin signaling in cells of the osteoclastic lineage and its possible effects on the regulation of bone mass is less known. Previous studies have shown that conditional ablation of βcatenin in osteoclasts induces a decrease in bone mass associated with an increase in osteoclasts, and this fact has been attributed to an increase in osteoclastogenesis. However, other alternative possibilities have not been evaluated, such as that a decrease in the normal osteoclast apoptosis may also contribute to the greater number of osteoclasts. In this paper, to obtain information about this fact, we generated mice in which βcatenin was selectively eliminated from cells of the monocyte/macrophage lineage using an allele flanked by βcatenin (Catnb.) together with the deletion line LisozimaMCre (LysMCre). The threeβdimensional analysis of the bones of the Catnbf/f;LysM mice revealed a significant decrease in the thickness of the femoral cortex, while the trabecular bone of the vertebrae was not affected. This phenotype was associated with a greater number of osteoclasts on the bone surface. The number of osteoclasts in the cultures from the Catnbf/f;LysM mice was twice as high as in the cultures obtained from the control mice. The administration of WNT3a attenuated the osteoclast formation induced by MβCSF and RANKL in vitro. In addition, WNT3a promoted apoptosis of osteoclasts, and this effect was counteracted, both by the presence of DKK1 and by the absence of βcatenin. Taken together, these results support a cellular autonomous effect of βcatenin in the osteoclast, and provide convincing evidence of the proapoptotic role of βcatenin in these cells.
Key words bone; osteoclasts; lysozyme M, β-catenin; Wnt; WNT3a
INTRODUCTION
The accumulated evidence over the past few years has established that the Wnt/β-catenin pathway is crucial for bone formation and the maintenance of skeletal homeostasis1,2. Wnt proteins exert their cellular functions by activating different signaling pathways, commonly called canonical pathway and non-canonical pathways3. The former acts by controlling the amount of β-catenin not associated with cadherin, while the other routes do not require the presence of β-catenin4. At present, the signaling pathway mediated by β-catenin is the best studied and understood. Activation of the Wnt/β-catenin pathway begins at the cell membrane with the binding of certain Wnt ligands, such as Wnt3a, to the transmembrane receptors of the Frizzled family. This binding recruits the LRP5/6 co-receptor (low-density-lipoprotein receptor-related protein 5/6), to form a ternary complex that destabilizes a cytoplasmic conglomerate of proteins that would otherwise phosphorylate the β-catenin of the cytoplasm for its destruction in the proteasome5-7. So, after ligand binding to the receptor, β-catenin is not phosphorylated or destroyed, and, therefore, can accumulate in the cytoplasm, from where it will be transferred to the nucleus. There it joins the transcription factor TCF/LEF (T-cell factor/lymphoid enhancer factor) and induces target gene expression8.
The initial demonstration of the canonical Wnt pathway involvement in osteogenesis was provided by the identification of loss-of-function (LOF) and gain-of-function (GOF) mutations of the LRP5 co-receptor, responsible for the osteoporosis-pseudoglioma syndrome and of the hereditary phenotype of high bone mass, respectively9,10. The bone phenotypes of these mutations could be reproduced in mouse models with the function of the genetically modified LRP5/6 receptor. These studies revealed the osteo-anabolic role of Wnt/β-catenin signaling11. More recently, studies carried out with mice in which β-catenin activity has been manipulated in cells of osteoblastic lineage have established that β-catenin increases bone mass through different mechanisms depending on the stage of differentiation in which the osteoblastic cell is found12-14. Thus, the GOF of β-catenin exclusively in precursor cells stimulates its proliferation and maturation, but suppresses the alternative fate towards chondrocytic differentiation, thus producing an increase in bone mass15. Furthermore, when the GOF of β-catenin is produced in a later stage of the differentiation, a high bone mass is also achieved, but this occurs at indirect mechanism rates, that is, β-catenin induces the expression of osteoprotegerin (OPG) in the osteoblast, and OPG attenuates osteoclastogenesis16,17.
The indirect participation of osteoclasts as mediators of some of the effects of Wnt/β-catenin signaling raised the question of whether this pathway could also have a direct role in the function of osteoclasts when activated in them. Otero et al.18 and Albers et al.19 eliminated β-catenin from osteoclast precursors using the mouse model LisozimaMCre (LysMCre), and found a decrease in bone mass in the trabecular compartment, with a parallel increase in the number of osteoclasts, thus showing that the β-catenin of osteoclasts is, in fact, involved in bone homeostasis. These authors, however, attributed the increased number of osteoclasts to an exclusive increase in osteoclastogenesis, without considering possible effects on apoptosis. We believe it is important to clarify this issue, since β-catenin is part of a signaling pathway that is currently considered an interesting therapeutic target. To obtain an idea of this problem, we have generated a similar mouse model, in which we have analyzed its phenotype, and also possible effects on osteoclast apoptosis.
MATERIAL AND METHODS
1. Reagents
The WNT3a protein, the Dickkopf 1 protein (DKK1), the macrophage colony stimulating factor (M-CSF) and the soluble receptor activator of nuclear factor kappa-B ligand recombinant proteins (sRANKL) were purchased from R & D Systems (Minneapolis, USA).
2. Mutant mice
All procedures with animals were carried out in accordance with European Union standards and 3R principles. The experiments were reviewed and approved by the Bioethics Committee of the University of Cantabria. The mouse with the β-catenin allele flanked by the flox20 sequence and the LysMCre21 line have previously been described and obtained from the Jackson Laboratory. Mice were genotyped by PCR, using genomic DNA extracted from tail biopsy. The primers used for the detection of LysMCre were the following; foward GCGGTCTGGCAGTAAAAACTATC and reverse: GTGAAACAGCATTGCTGTCACTT, and the product size 102 bp. Reverse: CACCATGTCCTCTGTCTATTC, and the product size in this case would be 324 bp. The experimental mice were generated by a mating strategy consisting of two steps. The LysMCre heterozygous mice were crossed with mice whose β-catenin gene was flanked by flox sequences in homozygosis (β-cateninf/f). To generate mice homozygous for the conditional allele of β-catenin with and without the Cre allele, the β-cateninf/+ mice;LysMCre crossed with the β-cateninf/f mice.
3. Micro-CT
A micro-CT analysis of the fifth lumbar vertebra was carried out after dissecting the bones, cleaning them, fixing them in ethanol, loading them in 12.3 mm diameter examination tubes and obtaining an image (μCT40, Scanco Medical, Basserdorf, Switzerland ). The scans were integrated into 3-D voxel images (1,024 x 1,024 pixel matrices for each individual planar stack) and a Gaussian filter (sigma=0.8, support=1) was used to reduce signal noise. A threshold of 200 was applied to all the analyzed scans. The scans were performed at a medium resolution (E=55 kVp, I=145 μA, integration time=200 ms). The entire vertebral body was scanned with a transverse orientation that excluded pedicles and joint processes. The manual analysis excluded the cortical bone from the analysis. All trabecular measurements were made by manually drawing contours every 10 to 20 cuts and using the voxel count for bone volume per tissue volume and spherical filling distance transformation rates without assumptions about bone shape as a rod or plate. trabecular microarchitecture. The cortical thickness was measured in the middle of the femoral diaphysis.
4. Bone histology
The lumbar vertebrae (L1, L2 and L3) and the left femur were fixed in 4% paraformaldehyde overnight at 4ºC and decalcified for eight to nine days in 9% EDTA (pH 7.4) before inclusion in paraffin. The histomorphometric examination was performed on longitudinal sections of 7 ?m of the femur for cortical bone and on the frontal sections of the vertebrae for spongy bone.
5. Cell cultures
To quantify the osteoclast progenitor cells, the bone marrow (BM) was purged from the long bones and seeded at a density of 50,000 cells/cm2 in 48-well plates. After remaining 4-5 days in the culture dish with α-MEM medium (Invitrogen), supplemented with 10% FBS, 1% PSG, 30 ng/ml M-CSF and 30 ng/ml sRANKL (R & R). D Systems), the osteoclasts were fixed with 10% formalin for 15 min and stained for tartrate-resistant acid phosphatase (TRAP). Osteoclasts were quantified taking into account multinuclear cells and positive for TRAP staining. For the rest of the cultures, purified bone marrow cells were used, ie not adhered to the plate. Macrophages and osteoclasts were developed from bone marrow (BM) cells not adhered to the culture dish, and cultured for 4 days in the presence of M-CSF (130 ng/ml) to obtain macrophages, or for 4 days in the presence of M-CSF (30 ng/ml) and 30 ng/ml of RANKL to obtain osteoclasts.
6. Real-time quantitative PCR (Quantitative real-time PCR, qRT-PCR)
The total RNA was extracted with TRIzol reagents (Life Technologies). 1 μg of total RNA was used to produce first strand cDNA using the enzyme m-MLV RT (Invitrogen). The qRT-PCR was carried out using PreMix Ex taq (Takara) and the data was analyzed using the Biorad software. The primers and probes for β-catenin [Mm01350385_g1 (fam)] and gapdh [Mm99999915_g1 (vic)] were manufactured by the TaqMan Gene Expression Assays service (Applied Biosystems). The relative levels of mRNA expression were normalized with the S2 ribosomal protein gene by the ΔCt method22.
7. Analysis by Western blot
The protein levels of β-catenin and β-actin were analyzed using a mouse monoclonal antibody that recognizes β-catenin (BD Biosciences), and a mouse monoclonal antibody that recognizes β-actin (Sigma-Aldrich).
8. Analysis of apoptosis
Osteoclasts derived from OM cells not adhered to the culture dish extracted from Catnbf/f;LysMCre mice and their controls were obtained. Once developed the osteoclasts in the culture dish were treated with WNT3a. After 24 hours of treatment, the cultures were fixed and stained by TUNEL and TRAP. The total number of osteoclasts and the number of apoptotic osteoclasts in each plate were quantified. They were considered apoptotic when at least one of their nuclei was TUNEL positive. The TUNEL method was carried out using the FragEL DNA fragmentation detection kit (EMD Chemicals, San Diego, California, USA) before staining for TRAP. Multinuclear TRAP positive and TUNEL positive cells were enumerated.
The activity of caspase-3 was measured by determining the degradation of the fluorometric substrate DEVD (Biomol Research Laboratories, Plymouth Meeting, Pennsylvania, USA), and the protein concentration was measured using a kit compatible with Bio-Rad detergent (Bio-Rad Laboratories, Hercules, California, USA).
RESULTS
1. The specific elimination of β-catenin from osteoclast precursors
Conditional inactivation of β-catenin (Catnb) in cells of osteoclastic lineage was performed by crossing mice harboring a β-catenin floxeado allele (Catnbf)20 with mice expressing the Cre recombinase enzyme under the control of the gene regulatory elements Lysozyme (LysMCre)21. This Cre line induces the recombination of the floxeado allele specifically in cells of monocyte-macrophage lineage and neutrophils. From this crossing, two cohorts (males and females) of animals were generated; an experimental cohort, with Catnbf/f;LysMCre genotype and another control with Catnbf/f genotype.
The Catnbf/f;LysMCre mice were born in the expected Mendelian proportion, with similar body weight (Figure 1A) to the controls of the same litter and showed no evident phenotype. The cleavage of the β-catenin gene was confirmed by qRT-PCR. The β-catenin mRNA levels were analyzed in ex vivo cultures of bone marrow (BM) derived macrophages and osteoclasts. The macrophages and osteoclasts obtained from Catnbf/f;LysMCre mice exhibited a 70% and 60% decrease in β-catenin expression, respectively (Figure 1B). We attribute the limited efficacy of recombination to the presence of cells from lineages other than myeloid in the culture plate that expressed normal levels of β-catenin. As expected, the expression level of β-catenin mRNA in kidney, liver and spleen was indistinguishable between the two genotypes (Figure 1C).

Bars: values expressed as mean ± standard deviation; Oc: osteoclast; *p≤0.05.
Figure 1 Catnbf/f;LysM-cre+/-mice express lower levels of β-catenin in macrophages and osteoclasts. A: total body weight of two cohorts (15 animals per group) of mice Catnbf/f;LysM-cre+/-and their control litter Catnbf/f of 28 weeks of age. B: quantitative analysis of mRNA by real-time PCR (Real Time-PCR) in macrophages and osteoclasts developed from non-adherent MO cells cultured in the presence of M-CSF for 4 days, and M-CSF plus RANKL for 5 days, respectively. C: quantitative analysis of soft tissue mRNA (indicated) obtained from 28-week-old mice (n=5)
2. Animals lacking β-catenin in osteoclast precursors have a lower cortical bone thickness
At 28 weeks of age, a cohort of 15 animals was sacrificed by sex and genotype and the bone architecture of the fifth lumbar vertebra and the right femur was examined by micro-CT. The analysis revealed a reduced thickness of the cortical bone (Figure 2A), both in the males and in the females Catnbf/f;LysMCre. However, the absence of β-catenin in the osteoclast precursors did not significantly alter the trabecular bone mass, neither in the vertebrae nor in the femur (Figure 2B). Neither the intertrabecular space, the trabecular thickness and the number of trabeculae (not shown) were affected. These results suggest that Wnt/?catenin signaling in osteoclasts is important for the maintenance of cortical bone mass.

Barras: valores expresados como media ± desviación estándar; Oc: osteoclasto; *p=0,05.
Figure 2 Los ratones Catnbf/f;LysM-cre+/- tienen menor masa ósea cortical. Mediciones de tomografía micro computarizada (μ-CT) realizadas en huesos de ratones de 28 meses (n=12-15 ratones por grupo). A: espesor cortical (Cortical th) determinado en fémures. B: BV/TV, volumen óseo por volumen total de tejido determinado en L5 y fémur derecho (solo se muestran las hembras)
3. Catnbf/f;LysMCre mice present more osteoclasts in the endosteum and more osteoclast progenitors in the bone marrow (BM)
Next, we wanted to quantify the number of osteoclasts present on the surface of the cortical bone. What we found was that, according to a decreased cortical thickness, the number of osteoclasts on the endocortical surface of the femur of the Catnbf/f;LysMCre mice was increased, as compared to the control animals (Figure 3A). The vertebral trabecular bone seemed to show the same tendency. However, the difference was not significant (p=0.06).

Barras: valores expresados como media ± desviación estándar; Oc: osteoclasto; *p=0,05.
Figure 3 The Catnbf/f; LysM-cre+/-mice have more osteoclasts than the control litter. A: histomorphometric analysis of decalcified longitudinal sections of the femur and L1-L3 vertebrae of female mice aged 28 months (n=5 mice per group). Photomicrographs (x40) show representative areas of the endosteal bone surface after TRAP staining. B: number of TRAPpositive cells developed from MO cells, obtained from the femurs of 28-week-old mice, and cultured in the presence of M-CSF and RANKL for 5 days. Cultures were performed in triplicate of each of three animals separately. Each bar represents triplicates of each mouse, n=3 per group. Photomicrographs (x40) show representative areas of the culture plate
To examine whether the greater number of osteoclasts in bone could be explained by an increase in osteoclastogenesis, the number of these progenitors in MO was quantified. For this purpose, the MO cells obtained from the long bones of a 28-week-old mice were cultured in the presence of RANKL and M-CSF for 5 days. Three mice were used per genotype and determinations were made in triplicate for each of them. The number of osteoclasts that developed in cultures from Catnbf/f;LysMCre mice showed a normal morphology (Figure 3B). However, the amount was twice as high as in the cultures from the control litter (426±18 per well vs 238±77, p=0.015) (Figure 3B). This result suggests that Wnt/β-catenin signaling in osteoclast precursors and their offspring attenuates the number of mature osteoclasts.
4. Proapoptotic effect of WNT3a requires the presence of β-catenin
To examine the cellular mechanisms through which Wnt/β-catenin decreased the number of osteoclasts, we first determined whether the addition of WNT3a to the culture plate of the osteoclast precursors interfered with the development thereof. As shown in Figure 4A, the presence of WNT3a in the culture medium decreased the number of osteoclasts induced by the presence of M-CSF and RANKL. In addition, this fact seemed to be dose dependent. Western blot analysis confirmed an increase in β-catenin levels in osteoclasts exposed to WNT3a (Figure 4B). In addition, treatment of the culture with DKK1, an inhibitor of the LRP5/6 co-receptor, prevented the increase of β-catenin induced by WNT3a. The set of these findings indicates that WNT3a inhibits the development of osteoclasts by stimulating the canonical Wnt pathway.
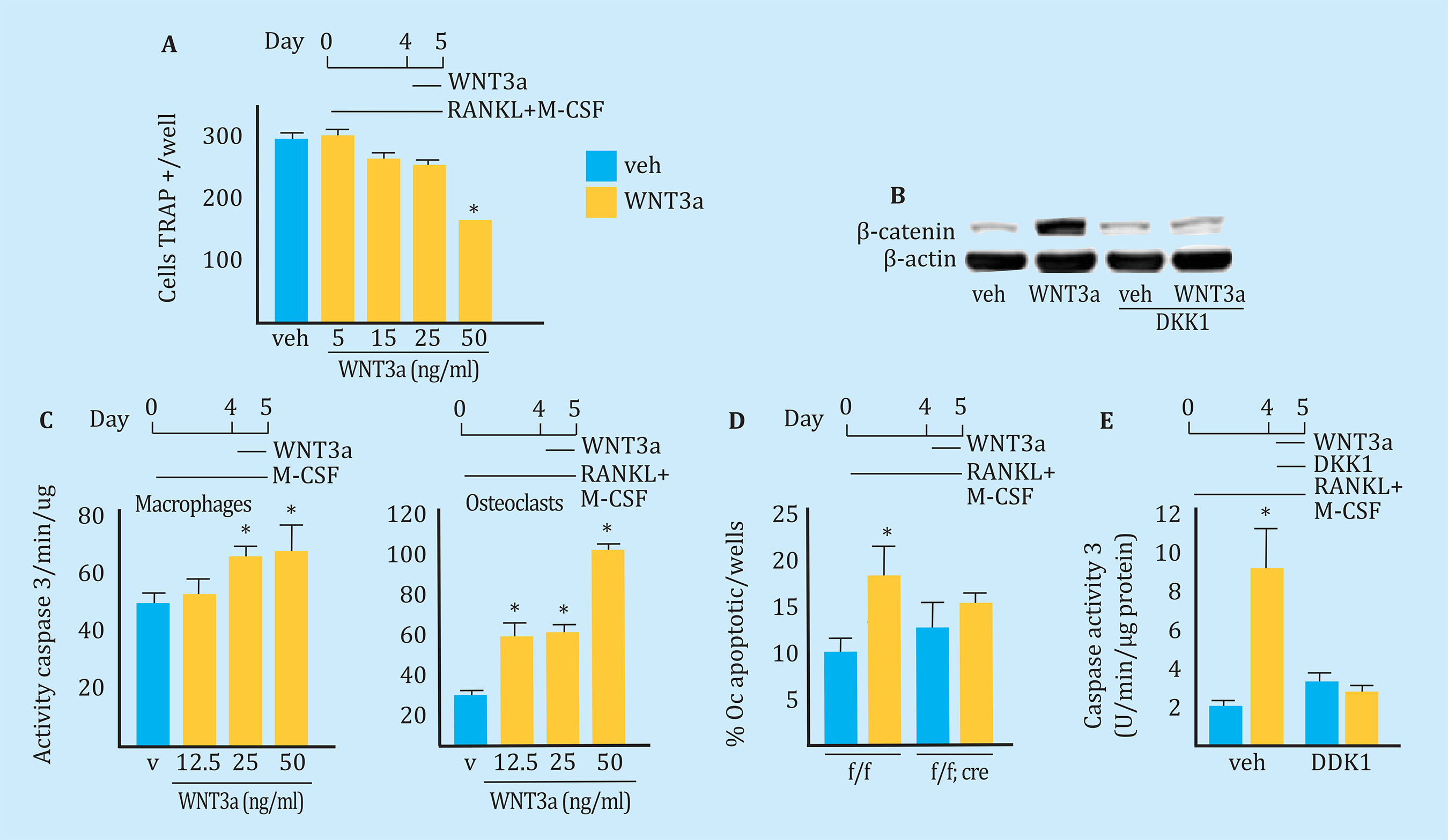
Bars: values expressed as mean ± standard deviation; Oc: osteoclast; *p=0.05.
Figure 4 WNT3a induces apoptosis of osteoclasts through the canonical Wnt pathway. A: number of TRAP-positive cells generated from MO cells not adhered to the plate obtained from C57BL/6 mice and cultured with M-CSF, RANKL and placebo (veh) or increasing doses of recombinant protein WNT3a as indicated. B: Western blot analysis of β-catenin in mature osteoclasts treated with veh, WNT3a, DKK1 or both. C: caspase 3 activity in mature macrophages and osteoclasts generated from non-adherent MO cells obtained from C57BL/6 mice and treated with veh or with different doses of WNT3a for 16 hours. D: TUNEL assay performed on mature osteoclast cultures generated from MO cells of Catnbf/f;LysM-cre mice cultured with M-CSF and RANKL for 5 days and treated with veh or WNT3a (50 ng/ml) for 24 hours. hours. E: activity of caspase 3 in mature osteoclasts generated from nonadherent MO cells of C57BL/6 mice, and treated with veh, WNT3a (50 ng/ml) or DKK1 (1 μg/ml) for 24 h. AFU, arbitrary fluorescent units
The half-life of osteoclasts is known to be very short and they die by apoptosis. Next, we examined the effect of Wnt signaling on the apoptosis of macrophages and osteoclasts. As shown in figure 4C, apoptosis was determined by measuring the activity of caspase-3 after administration of increasing doses of WNT3A to the macrophage and osteoclast cultures. The results showed that WNT3a induced apoptosis in both macrophages and osteoclasts. The proapoptotic effect of WNT3 was also dependent on the dose used. The effect of WNT3a on osteoclast apoptosis was also analyzed by TUNEL (Figure 4D), and, in the same way as observed in the previous experiment, we saw that treatment with WNT3a increased the percentage of TUNEL positive osteoclasts. However, the presence of WNT3 had no deleterious effect on the osteoclast cultures obtained from Catnbf/f;LysMCre mice (Figure 4D), lacking β-catenin. Likewise, the addition of DKK1 to the cultures abolished the proapoptotic actions of WNT3a (Figure 4D), indicating that the proapoptotic effect of WNT3a requires the presence of β-catenin. Taken together, these results support the hypothesis that the canonical signaling pathway of Wnt/β-catenin exerts proapoptotic effects on osteoclasts.
DISCUSSION
In this study, we have analyzed the bone characteristics of animals lacking β-catenin in cells of the monocyte/macrophage lineage, which are the precursors of osteoclasts. These animals show a reduced cortical thickness associated with a greater number of osteoclasts on the surface of the endosteum and a greater number of osteoclast progenitors in the BM. In addition, we demonstrate that the stimulation of Wnt/β-catenin signaling in osteoclasts attenuates the amount of developed osteoclasts induced by the presence of M-SCF and RANKL, and promotes their apoptosis.
Otero et al.18 and Albers et al.19 used the same LysMCre line to eliminate β-catenin from osteoclast precursors. Both groups described a decrease in bone mass in the trabecular compartment, with a parallel increase in the number of osteoclasts, which they attributed to an exclusive increase in osteoclastogenesis. However, they did not address the possibility that a decrease in osteoclastic apoptosis was also implicated. In our study, we demonstrated an increase in osteoclasts on the endocortical surface of the femoral bone, and an increased number of osteoclasts generated in cell cultures obtained from Catnbf/f;LysMCre animals, compared to controls. We also observed that the stimulation of Wnt/β-catenin signaling in the MO cells obtained from wild mice decreases the number of osteoclasts developed in the culture plate. Our results, therefore, coincide with those of Otero et al.18 and Albers et al.19. In addition, we have addressed the question of a possible involvement of osteoclastic apoptosis as part of the mechanism of action underlying the observed phenotypic findings. Our findings, in this sense, indicate that the decrease in the number of osteoclasts induced by the activation of Wnt/β-catenin is clearly due to the stimulation of apoptosis of macrophages and osteoclasts.
Several studies have shown that alterations in the survival of osteoclasts modify bone mass23-26. In fact, it is well established that estrogen protects the skeleton, in part, through proapoptotic effects on osteoclasts27,28. The elimination of estrogen receptor alpha in cells of the osteoclastic lineage, similar to the elimination of β-catenin, increases the number of osteoclasts and decreases bone mass. Unlike estrogen, glucocorticoids promote the loss of bone mass, at least in part, through the prolongation of the useful life of osteoclasts29.
Wnts proteins have a positive effect on the survival of osteoblasts and also on osteoblastic progenitors that have not yet been compromised30. This antiapoptotic action of Wnts proteins on osteoblasts has been postulated as one of the mechanisms by which Wnt signaling increases bone mass31. Although the Wnt/β-catenin pathway is best known for its pro-survival effects, it can also exert proapoptotic actions. For example, the apoptosis of rat cardiomyoblasts induced by reoxygenation after hypoxia is regulated by WNT3a, through a mechanism dependent on β-catenin32. In addition, in line with these findings, Wnt/β-catenin signaling decreases the cellular invasiveness of melanoma33, enhancing the expression of proapoptotic proteins, such as BIM and PUMA, and decreasing levels of antiapoptotic proteins, such as MCL34.
In our study, through the analysis of caspase activity, or TUNEL assays, we have found that WNT3a induces osteoclast apoptosis. This effect is contrary to its previously mentioned pro-survival effect on osteoblasts30. Interestingly, like Wnts, estrogens and glucocorticoids exert opposite effects on the apoptosis of osteoblasts and osteoclasts29,35,36.
To conclude, our findings suggest that the inhibitory effects of β-catenin on osteoclasts may be attributed to proapoptotic effects and support the claim that the osteoprotective effects of the canonical Wnt pathway also result from direct action through the osteoclastic lineage cells. Therefore, Wnt/β-catenin signaling in the bone environment has an osteoprotective effect exerted through both the osteoblastic and osteoclastic lineages.
Bibliografía
1 Hartmann C. A Wnt canon orchestrating osteoblastogenesis. Trends Cell Biol. 2006;16(3):151-8. [ Links ]
2 Baron R, Kneissel M. WNT signaling in bone homeostasis and disease: from human mutations to treatments. Nat Med. 2013;19(2):179-92. [ Links ]
3 Eisenmann DM. Wnt signaling. Worm-Book. 2005 Jun 25:1-17. [ Links ]
4 Gómez-Orte E, Sáenz-Narciso B, Moreno S, Cabello J. Multiple functions of the noncanonical Wnt pathway. Trends Genet. 2013;29(9):545-53. [ Links ]
5 Tamai K, Semenov M, Kato Y, Spokony R, Liu C, Katsuyama Y, et al. LDL-receptorrelated proteins in Wnt signal transduction. Nature. 2000;407(6803):530-5. [ Links ]
6 He X, Semenov M, Tamai K, Zeng X. LDL receptor-related proteins 5 and 6 in Wnt/beta-catenin signaling: arrows point the way. Development. 2004; 131:1663-77. [ Links ]
7 Liu C, Li Y, Semenov M, Han C, Baeg GH, Tan Y, et al. Control of beta-catenin phosphorylation/degradation by a dual-kinase mechanism. Cell. 2002; 108(6):837-47. [ Links ]
8 Bienz M, Clevers H. Armadillo/betacatenin signals in the nucleus--proof beyond a reasonable doubt? Nat Cell Biol. 2003;5(3):179-82. [ Links ]
9 Gong Y, Slee RB, Fukai N, Rawadi G, Roman-Roman S, Reginato AM, et al. Osteoporosis-Pseudoglioma Syndrome Collaborative Group. LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Cell. 2001;107(4):513-23. [ Links ]
10 Boyden LM, Mao J, Belsky J, Mitzner L, Farhi A, Mitnick MA, et al. High bone density due to a mutation in LDL-receptor-related protein 5. N Engl J Med. 2002;346:1513-21. [ Links ]
11 Zylstra CR, Wan C, VanKoevering KK, Sanders AK, Lindvall C, Clemens TL, et al. Gene targeting approaches in mice: assessing the roles of LRP5 and LRP6 in osteoblasts. J Musculoskelet Neuronal Interact. 2008;8(4):291-3. [ Links ]
12 Hu H, Hilton MJ, Tu X, Yu K, Ornitz DM, Long F. Sequential roles of Hedgehog and Wnt signaling in osteoblast development. Development. 2005;132:49-60. [ Links ]
13 Day TF, Guo X, Garrett-Beal L, Yang Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev Cell. 2005;8:739-50. [ Links ]
14 Hill TP, Spater D, Taketo MM, Birchmeier W, Hartmann C. Canonical Wnt/ beta-catenin signaling prevents osteoblasts from differentiating into chondrocytes. Dev Cell. 2005;8:727-38. [ Links ]
15 Rodda SJ, McMahon AP. Distinct roles for Hedgehog and canonical Wnt signaling in specification, differentiation and maintenance of osteoblast progenitors. Development. 2006;133:3231-44. [ Links ]
16 Glass DA, Bialek P, Ahn JD, Starbuck M, Patel MS, Clevers H, et al. Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev Cell. 2005;8:751-64. [ Links ]
17 Kramer I, Halleux C, Keller H, Pegurri M, Gooi JH, Weber PB, et al. Mol Osteocyte Wnt/beta-catenin signaling is required for normal bone homeostasis. Cell Biol. 2010;30(12):3071-85. [ Links ]
18 Otero K, Shinohara M, Zhao H, Cella M, Gilfillan S, Colucci A, et al. TREM2 and ?-catenin regulate bone homeostasis by controlling the rate of osteoclastogenesis. J Immunol. 2012;188(6):2612-21. [ Links ]
19 Albers J, Keller J, Baranowsky A, Beil FT, Catala-Lehnen P, Schulze J, et al. Canonical Wnt signaling inhibits osteoclastogenesis independent of osteoprotegerin. J Cell Biol. 2013;200(4):537-49. [ Links ]
20 Brault V, Moore R, Kutsch S, Ishibashi M, Rowitch DH, McMahon AP, et al. Inactivation of the beta-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development. 2001;128(8):1253-64. [ Links ]
21 Clausen BE, Burkhardt C, Reith W, Renkawitz R, Förster I. Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic Res. 1999;8(4):265-77. [ Links ]
22 Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) Method. Methods. 2001;25(4):402-8. [ Links ]
23 Jilka RL. Biology of the basic multicellular unit and the pathophysiology of osteoporosis. Med Pediatr Oncol. 2003;41(3):182-5. [ Links ]
24 Weinstein RS, Manolagas SC. Apoptosis and osteoporosis. Am J Med. 2000; 108(2):153-64. [ Links ]
25 Manolagas SC. Birth and death of bone cells: basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr Rev. 2000;21(2):115-37. [ Links ]
26 Xing L, Boyce BF. Regulation of apoptosis in osteoclasts and osteoblastic cells. Biochem Biophys Res Commun. 2005;328(3):709-20. [ Links ]
27 Nakamura T, Imai Y, Matsumoto T, Sato S, Takeuchi K, Igarashi K, et al. Estrogen prevents bone loss via estrogen receptor alpha and induction of Fas ligand in osteoclasts. Cell. 2007;130(5):811-23. [ Links ]
28 Martin-Millan M, Almeida M, Ambrogini E, Han L, Zhao H, Weinstein RS, et al. The estrogen receptor-alpha in osteoclasts mediates the protective effects of estrogens on cancellous but not cortical bone. Mol Endocrinol. 2010;24(2):323-34. [ Links ]
29 Jia D, O'Brien CA, Stewart SA, Manolagas SC, Weinstein RS. Glucocorticoids act directly on osteoclasts to increase their life span and reduce bone density. Endocrinology. 2006;147(12):5592-9. [ Links ]
30 Almeida M, Han L, Bellido T, Manolagas SC, Kousteni S. Wnt proteins prevent apoptosis of both uncommitted osteoblast progenitors and differentiated osteoblasts by beta-catenin-dependent and -independent signaling cascades involving Src/ERK and phosphatidylinositol 3-kinase/AKT. J Biol Chem. 2005;280(50):41342-51. [ Links ]
31 Krishnan V, Bryant HU, Macdougald OA. Regulation of bone mass by Wnt signaling. J Clin Invest. 2006;116(5):1202-9. [ Links ]
32 Zhang Z, Deb A, Zhang Z, Pachori A, He W, Guo J, et al. Secreted frizzled related protein 2 protects cells from apoptosis by blocking the effect of canonical Wnt3a. J Mol Cell Cardiol. 2009;46(3): 370-7. [ Links ]
33 Smalley KS, Contractor R, Haass NK, Kulp AN, Atilla-Gokcumen GE, Williams DS, et al. An organometallic protein kinase inhibitor pharmacologically activates p53 and induces apoptosis in human melanoma cells. Cancer Res. 2007;67(1):209-17. [ Links ]
34 Zimmerman ZF, Kulikauskas RM, Bomsztyk K, Moon RT, Chien AJ. Activation of Wnt/β-catenin signaling increases apoptosis in melanoma cells treated with trail. PLoS One. 2013;8(7):e69593. [ Links ]
35 Manolagas SC, O'Brien CA, Almeida M. The role of estrogen and androgen receptors in bone health and disease. Nat Rev Endocrinol. 2013;9(12):699-712. [ Links ]
36 O'Brien CA, Jia D, Plotkin LI, Bellido T, Powers CC, Stewart SA, et al. Glucocorticoids act directly on osteoblasts and osteocytes to induce their apoptosis and reduce bone formation and strength. Endocrinology. 2004;145(4):1835-41. [ Links ]
Received: July 19, 2018; Accepted: February 19, 2019
 Este es un artículo publicado en acceso (Open Access) abierto bajo la licencia Creative Commons Attribution Non-Commercial, que permite su uso, distribución y reproducción en cualquier medio, sin restricciones siempre que sin fines comerciales y que el trabajo original sea debidamente citado.
Este es un artículo publicado en acceso (Open Access) abierto bajo la licencia Creative Commons Attribution Non-Commercial, que permite su uso, distribución y reproducción en cualquier medio, sin restricciones siempre que sin fines comerciales y que el trabajo original sea debidamente citado.
