










 Servicios personalizados
Servicios personalizados
 texto en
texto en  Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
La osteoporosis (OP) es una de las enfermedades metabólicas más frecuentes en el mundo. Se caracteriza por la pérdida de masa ósea y deterioro de la microarquitectura del hueso, predisponiendo a los pacientes a sufrir fracturas por fragilidad (1). Esta enfermedad es considerada como una condición subclínica hasta que se complica con una fractura, lo que supone una carga médica y personal, además de que también implica un alto costo socioeconómico y recursos necesarios para la atención de individuos que la padecen (2,3). La OP se considera como una enfermedad que afecta típicamente a mujeres posmenopáusicas. Sin embargo, en los últimos años se ha observado que un tercio de fracturas de fémur ocurren en hombres y la incidencia de fracturas vertebrales puede superar más de la mitad de la descritas en mujeres (4,5). Actualmente se estima que 75 millones de personas en Europa, Estados Unidos y Japón son afectados por OP, siendo la causa de hasta 8,9 millones de fracturas por fragilidad ósea. En México, en 2010, de acuerdo con el censo de población y vivienda, se reportó que la población total fue 112 millones de habitantes de los cuales el 17 % correspondía a la población de adultos mayores de 50 años. Dentro de esta población el 17 % de las mujeres y el 9 % de los hombres mexicanos presentaron OP en columna lumbar, mientras que el 16 % de mujeres y el 6 % de hombres mexicanos presentaron OP en cadera, respectivamente (5,6). Actualmente, de acuerdo con datos del censo de población y vivienda 2020 la población mexicana se compone de 126 millones de habitantes que se encuentran enfrentando una transición epidemiológica con un incremento en la esperanza de vida, donde la población mayor de 50 años corresponde al 17,46 % y de esta población se estima que 10 millones de individuos viven con OP, lo que implica que una de cada tres mujeres por uno de cada cinco hombres presentará este padecimiento (7,8). De acuerdo con estas estadísticas, la OP en hombres se considera como una creciente preocupación de salud pública lo que ha promovido el desarrollo de guías clínicas para el cuidado de esta enfermedad que ahora abordan la evaluación y el tratamiento en pacientes masculinos. Sin embargo, a pesar del desarrollo de estas guías, la OP masculina sigue considerándose como una afección subdiagnosticada y subtratada (9). Aunque el cuadro clínico entre hombres y mujeres es similar, existen algunas características particulares en la OP masculina, por ejemplo: en la mayoría de los casos el tipo de OP es “secundaria”, es decir que se origina por consecuencia de otras enfermedades, uso de fármacos o cambios en el estilo de vida, los criterios de densitometría para el diagnóstico no están bien validados, existe carencia de estudios que analicen el efecto de diferentes tratamientos en la prevención de los pacientes, no existen antecedentes de traumatismos o sobre el origen de las fracturas y, en el caso de los hombres, estos son menos propensos a caídas, además de que la esperanza de vida es más corta, lo que hace que las acciones terapéuticas empleadas en hombres sean diferentes a las empleadas en mujeres (10). Por lo tanto, el objetivo de este trabajo es realizar una revisión narrativa sobre el papel de las hormonas sexuales, su influencia sobre la densidad mineral ósea y su papel en el desarrollo de la OP masculina.
DESARROLLO DEL HUESO
El hueso es un tipo de tejido conectivo altamente especializado cuya función principal es proporcionar un soporte mecánico para actividad muscular y protección física de los tejidos y órganos internos, además de mantener la homeostasis y ser un depósito mineral a nivel sistémico (11). A lo largo del desarrollo existen diferencias importantes entre hombres y mujeres durante las primeras etapas de la vida, el desarrollo óseo está influenciado por diferentes factores como las hormonas sexuales, el nivel de actividad física y el tamaño corporal, y es en la adolescencia en la etapa que posiblemente tenga mayor influencia sobre la formación y desarrollo del sistema esquelético tanto en hombres como en mujeres. Durante la pubertad los hombres inician esta etapa de la vida más tarde que las mujeres y dura más tiempo, lo que podría causar diferencias en el desarrollo óseo entre ambos géneros. Un ejemplo de esto es que los hombres tienden a tener piernas más largas que las mujeres debido a que la fusión epifisiaria ocurre más tarde en hombres debido a que presentan un periodo de maduración ósea más largo (12). Por otro lado, las hormonas sexuales también ejercen un efecto sobre el crecimiento, la homeostasis del hueso y el final de la maduración ósea. En los hombres la testosterona juega un papel clave en el desarrollo de esqueletos más grandes, mientras que los estrógenos se han asociado con la reducción de la reabsorción del hueso, conservando así la masa ósea. Sin embargo, se ha reportado que la testosterona también tiene un efecto en la mejora de la formación ósea y se puede asociar con la reducción de la reabsorción del tejido óseo, pero esto puede deberse a la conversión de testosterona a estrógenos, lo que sugiere que los estrógenos podrían estar encargados de conservar la masa ósea y la testosterona de aumentar la masa ósea. Estas funciones de las hormonas sexuales en el desarrollo del hombre le confieren varias ventajas hasta llegar a la vida adulta ya que permiten proteger los huesos de fracturas por fragilidad con respecto a mujeres, pueden presentar un pico de masa ósea mucho más alto, un tamaño óseo más grande y una mayor fortaleza ósea (13).
ESTRÓGENOS EN EL DESARROLLO DEL HUESO
Los estrógenos son una familia de hormonas esteroideas que se componen por estradiol, estriol y estetrol. El estradiol (E2) es el estrógeno más frecuente y activo, se produce principalmente por los ovarios, aunque el tejido adiposo, los testículos, la corteza suprarrenal y el hígado también contribuyen a su producción. Los estrógenos se han asociado con el mantenimiento de la masa ósea y a través de observaciones clínicas; se ha establecido que la deficiencia de estrógenos sobre la masa ósea también es causa de OP en hombres, lo que sugiere su papel universal en el metabolismo del hueso (14). Los efectos de los estrógenos sobre la masa ósea se atribuyen principalmente a su acción de inhibir la reabsorción ósea que se lleva a cabo a través de los osteoclastos. Diversos estudios in vitro que utilizan líneas celulares de osteoblastos y células estromales, muestran que los estrógenos disminuyen la producción de citocinas osteoclastogénicas y aumentan la expresión de factores que inhiben la osteoclastogénesis (15,16). Por otro lado, el efecto que pueden tener los estrógenos sobre la formación ósea sigue siendo poco claro, ya que las células estromales y los osteoblastos expresan el receptor de estrógenos alfa (REα) y beta (REβ) los cuales pueden afectar a la diferenciación y a la formación ósea. Los osteoblastos se derivan de células stem mesenquimales (CSM) que también pueden dar origen a los adipocitos, por lo que es posible que exista un mecanismo de regulación que determine el linaje entre osteoblastos/adipocitos y sea un componente crítico en la vía reguladora de la osteoblastogénesis (17). Se ha observado que el aumento de la concentración de lípidos en medula ósea se asocia con la pérdida de masa ósea asociada con la edad, lo que implica una relación inversa entre la adipogénesis y la osteoblastogénesis (18). Además, se ha observado que la osteopenia inducida por ovariectomía está asociada con un aumento de la adipogénesis, por lo que en diferentes estudios in vitro, se ha planteado la hipótesis de que los estrógenos regulan negativamente la expresión de la lipoproteína lipasa (LPL), un marcador característico de la diferenciación de adipocitos, por lo que los estrógenos podrían regular la formación ósea a través de la inactivación de las células estromales de medula ósea de origen mesenquimal para inducir un cambio del linaje y dirigirlo hacia el osteoblasto (19). Por otro lado, se cree que la acción principal de los estrógenos es inhibir la reabsorción ósea derivada de la actividad de los osteoclastos. Los osteoclastos son células gigantes multinucleadas cuya función principal es desintegrar la matriz mineral ósea en la fase de reabsorción del remodelado óseo (20). El reclutamiento de precursores de osteoclastos, la diferenciación y la actividad de reabsorción están controlados por factores locales, por ejemplo: la vitamina D, las prostaglandinas, el TGF-β, IL-1, IL-6 y el TNF-α que estimulan la diferenciación y actividad de los osteoclastos a través de mecanismos directos o indirectos, mientras que la calcitonina inhibe su actividad. El destino de los osteoclastos después de la reabsorción ósea es desconocido; se sabe que factores como la calcitonina inactivan los osteoclastos sin inducir la muerte celular, mientras que factores como los bifosfonatos y la vitamina K2 inducen la muerte celular de los osteoclastos. Por otro lado, se cree que la actividad de los estrógenos sobre los osteoclastos está regulada indirectamente a través de células no osteoclásticas. Se ha observado que la disminución de estrógenos durante la menopausia o debido a un proceso de ovariectomía se asocia con un incremento en la expresión de IL-1, IL-6 y TNF-α y una disminución de TGF-β por parte de los monocitos de sangre periférica, células del estroma de medula ósea y osteoblastos (21). Otros factores también se encuentran involucrados en la diferenciación y activación de los osteoclastos; una de las vías clave es la señalización del receptor activador nuclear kappa B (RANK) que se expresa en los osteoclastos y se activa cuando se une al ligando del receptor activador nuclear kappa B (RANKL); en este mecanismo también puede intervenir la osteoprotegerina (OPG), una proteína que sirve como señuelo de RANKL, suprimiendo la activación de la diferenciación de osteoclastos. Los estrógenos son capaces de regular a RANKL y promover la expresión de OPG, reduciendo la reabsorción ósea a través de cambios en la expresión de proteínas celulares de osteoblastos humanos entre los que encontramos algunos miembros de la vía de señalización Wnt/β-catenina, que regulan negativamente la osteoclastogénesis y son mediadores de efectos anabólicos en el hueso (22,23). Además, los estrógenos inhiben la diferenciación y favorece la apoptosis de osteoclastos al aumentar la producción de TGF-β, mientras que en ausencia de estrógenos se induce la expresión de RANKL, conduciendo a la activación de la osteoclastogénesis (24) (Fig. 1A).
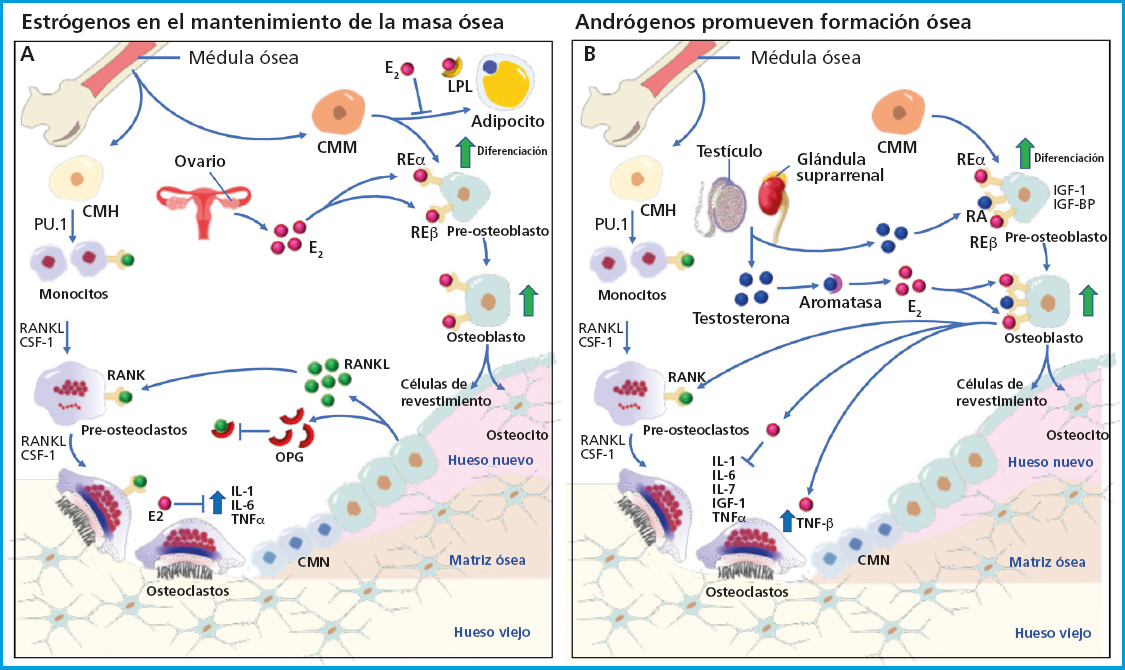
Figura 1. Representación esquemática sobre las unidades básicas multicelulares (BMU), su asociación con el remodelado óseo y el papel de las hormonas sexuales en el manteamiento y formación del tejido óseo. La activación del remodelado óseo inicia con la diferenciación de células madre hematopoyéticas a osteoclastos maduros capaces de reabsorber el tejido óseo, donde las citocinas CSF-1 y RANKL son requeridas en todas las etapas de diferenciación. Una vez terminada la reabsorción ósea, existe una atracción de células mononucleares hacia las unidades de remodelado óseo para reclutar precursores de osteoblastos que posteriormente se diferenciarán a osteoblastos maduros y osteocitos los cuales quedarán atrapados dentro de la matriz mineralizada y cuya función es de quimiotaxis y mecanosensora. Los osteoblastos son células formadoras de tejido óseo que se derivan de células madre mesenquimales y que requieren de la actividad de distintos factores para su diferenciación como la esclerotina, el factor de transcripción RUNX2 y la activación de las proteínas IGF-1 e IGF-BP. A. Se muestra la producción de estrógenos por parte de los ovarios, los cuales tienen la capacidad de inhibir la actividad de citocinas IL-1, IL6 y TNF-α, que son necesarias durante la osteoclastogénesis, además los estrógenos son capaces de unirse a los REα y REβ presentes en los precursores de osteoblastos para promover su diferenciación y en osteoblastos maduros para secretar citocinas inhibidoras de la osteoclastogénesis como la osteoprotegerina (OPG) la cual funciona como anzuelo de RANKL evitando su unión con los osteoclastos y ejerciendo una función de mantenimiento de la masa ósea. B. Se observa la actividad de la testosterona secretada por testículos y la glándula suprarrenal que se une a receptores de andrógenos presentes en los osteoblastos y promueve la diferenciación a osteoblastos maduros. La testosterona también actúa como inhibidor de la IL-6 que es necesaria para la osteoclastogénesis y a través de la mutasa es convertida a estrógenos, promoviendo la diferenciación osteoblástica, además de regular la expresión de citocinas necesarias en la osteoclastogénesis, por lo que la testosterona se puede promover la formación del tejido óseo.
ANDRÓGENOS EN EL DESARROLLO DEL HUESO
El término “andrógeno” hace referencia a la testosterona y sus precursores derivados del colesterol. La testosterona es un tipo de andrógeno predominante en hombres, que es secretado en un 95 % por los testículos y un 5 % por glándulas suprarrenales a través de la conversión de dehidroepiandrosterona (25). La testosterona se une a la albúmina y a la globulina transportadora de hormonas sexuales para que posteriormente sus efectos en el cuerpo sean regulados por su conversión local a 5α-dihidrotestosterona a través de tejidos periféricos, los cuales presentan alta afinidad debido a la gran cantidad de receptores de andrógenos (AR) que poseen. La testosterona ejerce fuertes efectos anabólicos y androgénicos que afectan a hombres y mujeres, influyendo significativamente en el crecimiento y mantenimiento de los huesos. En un estudio se demostró que, a través de la administración de testosterona en modelos murinos, estos presentaron un aumento en el ancho de la placa del crecimiento epifisario, los efectos observados en este trabajo fueron independientes de la hormona del crecimiento y del factor de crecimiento similar a insulina-1 (IGF-1); de este modo se mostraron los efectos de la testosterona sobre el desarrollo del hueso (26). En este sentido se ha demostrado que la testosterona juega un papel importante en el mantenimiento de la DMO de hombres de edad avanzada (27). Sin embargo, los niveles de testosterona en suero de hombres mayores, disminuye 1 % por año, lo que puede generar la aparición de síntomas clínicos como el síndrome de hipogonadismo de inicio tardío (LOH), el cual se caracteriza por depresión, irritabilidad, disfunción sexual, disminución de masa magra y disminución de la DMO, síntomas que podrían estar asociados con el envejecimiento, por lo que se ha propuesto la terapia de remplazo de testosterona como un tratamiento para mejorar la calidad de vida de hombres mayores que padecen LOH (28). Como se mencionó anteriormente, los estrógenos son necesarios para el mantenimiento de la DMO y en el caso de las mujeres disminuyen considerablemente durante la menopausia, mientras que en los hombres la testosterona disminuye lentamente con la edad, lo que permite mantener la DMO estable durante un tiempo prolongado. Por lo tanto, la OP es más frecuente en mujeres posmenopáusicas que en hombres mayores (29). En el metabolismo óseo la testosterona juega un papel importante ya que se convierte en dihidrotestosterona (DHT) altamente activa a través de la 5-reductasa en el citoplasma de las células diana; esto permite su unión con el receptor de andrógenos (RA) induciendo actividad androgénica. Además, la testosterona también se convierte en E2 debido a la actividad de la aromatasa, lo que permite su unión con los subtipos REα y REβ los cuales se asocian con el metabolismo del hueso. Los RA están presentes en condrocitos y osteoblastos en los que su nivel de expresión varía de acuerdo con la edad del individuo y los sitios óseos. La unión entre la testosterona y los RA en osteoblastos promueven la formación de hueso a través de la activación indirecta de citocinas y factores de crecimiento. Los osteoblastos sintetizan diversas citocinas que promueven la resorción del hueso, tales como la IL-6 o el TNF (30). Además, los andrógenos regulan positivamente el TGF-β y los factores de crecimiento IGF que estimulan la formación del hueso (31). Se ha observado que la deficiencia de testosterona promueve la expresión de RANKL a partir de osteoblastos y en consecuencia activa la diferenciación de osteoclastos aumentando la reabsorción ósea, lo que se ve reflejado en una disminución de la DMO. La diferenciación y proliferación de condrocitos y osteoblastos es inducida por la unión de IGF-1 con la proteína de unión a IGF (IGF-BP) que junto con la supresión de la apoptosis de condrocitos promueven la formación de hueso. Por lo tanto, la testosterona regula positivamente la expresión de IGF-1 e IGF-BP en osteoblastos (32). La testosterona también puede regular la osteoclastogénesis a través de la supresión de la actividad de la interleucina (IL) 6, cuya función es activar a los osteoclastos e inducir la reabsorción ósea. Por lo tanto, la disminución de los niveles de testosterona afecta negativamente los niveles de DMO. Sorprendentemente los niveles más altos de expresión de RA se han observado en osteocitos, que además de ser las células más abundantes dentro del hueso, se ha demostrado que producen diversos mediadores que pueden influir en la osteoclastogénesis, como el óxido nítrico, TGF-β, prostaglandinas o RANKL. La deficiencia de estrógenos y andrógenos conduce a una mayor prevalencia de apoptosis de los osteocitos (33). La apoptosis de los osteocitos puede estimular indirectamente la osteoclastogénesis al inducir que las células estromales/osteoblásticas secreten RANKL. Además, los osteocitos secretan OPG, que compite con RANK por su receptor en los osteoclastos. En los osteocitos, como en los osteoblastos, la secreción de OPG está regulada por la vía Wnt/β-catenina y se ha observado que, ratones que carecen de β-catenina en los osteocitos, son osteoporóticos debido al aumento del número de osteoclastos (34). Un mecanismo similar al que sucede en humanos. Los osteocitos controlan el proceso de remodelación ósea mediante la regulación directa e indirecta de la diferenciación y función de osteoclastos y osteoblastos, una alteración en el proceso conduce a la aparición de osteoporosis. En este sentido los subtipos REα y REβ juegan un papel importante en el mantenimiento de la DMO en hombres debido a que los estrógenos ejercen un mayor efecto que los andrógenos en la inhibición de la reabsorción ósea. Se ha observado que la pérdida de la función de REα y la deficiencia de aromatasa en pacientes masculinos induce el desarrollo de un fenotipo con una DMO extremadamente baja, por lo que se ha propuesto la terapia de remplazo de estrógenos como una opción para mejorar los niveles de DMO en pacientes adultos masculinos (35). El E2 generalmente se encarga de regular la apoptosis y la función de osteoclastos al aumentar la expresión del factor de crecimiento tumoral β (TNF-β). La expresión de IL-1, IL-6, IL-7, IGF-1, factor nuclear κB (NF-κB), RANK y el factor de necrosis tumoral α (TNF-α) aumentan, reduciendo la proliferación y actividad de los osteoblastos; es conocido que estos genes son blancos del efecto antirresortivo de los estrógenos sobre el hueso (36) (Fig. 1B).
RELACIÓN ENTRE LAS HORMONAS SEXUALES Y LA FRACTURA ÓSEA
Durante las actividades de la vida diaria las caídas y fracturas son comunes en hombres mayores. La búsqueda de herramientas que ayuden a la prevención de fracturas por fragilidad se ha convertido en uno de los principales objetivos a nivel mundial. La aparición de fracturas asociadas con el envejecimiento se debe principalmente a la disminución de la función física como la pérdida de la masa magra, debilidad muscular, fragilidad de los huesos, sarcopenia y la disminución de la DMO. Estudios recientes han identificado la relación entre la testosterona y el riesgo de fracturas, donde se ha observado que pacientes masculinos de edad avanzada con fracturas osteoporóticas presentaron niveles de testosterona muy bajos en comparación con grupos controles de la misma edad y origen étnico (37,38). Algunos estudios respaldan la hipótesis de que la deficiencia de testosterona se asocia con un aumento en la incidencia de caídas mientras que otros rechazan esta hipótesis (39). Las fracturas óseas más predominantes asociadas con disminución de la DMO debido a bajos niveles de testosterona, también pueden deberse a la relación de la testosterona con la fuerza muscular y el rendimiento físico en hombres, lo que podría conducir al desarrollo de sarcopenia y a un aumento en el riesgo de caídas. Actualmente se ha establecido que la relación entre la deficiencia de testosterona y la baja DMO es mucho más fuerte en adultos jóvenes masculinos con hipogonadismo de moderado a severo (40). Sin embargo, son pocos los estudios disponibles sobre la epidemiología de la OP masculina; esto puede deberse a los pequeños tamaños de muestra y posibles sesgos que presentan estos trabajos. Estudios de casos y controles donde se compara la prevalencia de hipogonadismo entre sujetos con OP y grupos controles han mostrado que las fracturas por OP son más frecuentes en pacientes con hipogonadismo con respecto a pacientes sin esta condición (41). Otros estudios han documentado un marcado incremento en el riesgo de sufrir fracturas por fragilidad entre pacientes con niveles bajos de testosterona y E2. Estos bajos niveles de hormonas sexuales se asocian con atrofia muscular y a la disminución de la masa magra total, por lo que es lógico pensar que una pérdida en la función muscular puede dañar el mecanismo defensivo contra las caídas, conduciendo a un aumento en la incidencia de fracturas en pacientes masculinos. Hoy en día es ampliamente aceptado que los trastornos del metabolismo óseo en pacientes con niveles bajos de estradiol pueden aumentar el riesgo de fracturas, lo que podría ser causado por un déficit de transformación de testosterona a estradiol debido a una disfunción de la enzima aromatasa, incluso algunos estudios han reportado el desarrollo de OP masculina severa debido a mutaciones en el receptor de estrógeno de la enzima aromatasa (42).
VÍAS DE SEÑALIZACIÓN EN EL METABOLISMO ÓSEO ACTIVADAS POR HORMONAS SEXUALES
Los E2 y otras hormonas esteroides son capaces de inducir la activación de diferentes vías de señalización uniéndose a su receptor a través de tres mecanismos: a) la señalización clásica, donde el E2 se une a los REα y REβ en el compartimiento citoplasmático, posteriormente este complejo se desplaza al núcleo donde forma homo- o heterodímeros que se unirán directamente a una secuencia de DNA especifica llamada elementos de respuesta a estrógenos (ERE); b) señalización independiente de ERE, donde el complejo E2/RE se desplaza hacia el núcleo e interactúa con factores de transcripción para secuestrarlos y modificar su interacción con el DNA, lo que conduce a una alteración en la expresión genética; y c) señalización no genotrópica (no implica cambios en la expresión genética), en el que el E2 envía señales a través de un receptor acoplado a proteína G (GPCR) en la membrana plasmática. Los RE se expresan en gran medida en el hueso y sus efectos se han atribuido a la actividad mediada por este receptor. Estos efectos fueron demostrados a través de un estudio donde a un grupo de ratones hembra ovariectomizados (OVX) REα-/- y un grupo de ratones macho orquiectomizados (ORX) REα-/- no respondieron a tratamiento con estrógeno exógeno. Los ratones REα-/- mostraron un aumento de aproximadamente 10 veces el nivel de E2 y 5 veces los niveles de testosterona, así como los niveles de IGF-1 alterados lo que condujo a una mayor actividad de osteoclastos y por lo tanto al desarrollo de un fenotipo osteoporótico (43). Existen REα en el núcleo y en la membrana celular, los cuales activan vías de señalización independientes de la transcripción que son activadas por mecanismos no genómicos del REα, donde el estrógeno ejerce efectos antioxidantes de manera independiente. El efecto biológico de la osteogénesis se relaciona con vías de señalización celular altamente específicas, entre las que se encuentra la vía de señalización de la fosfatidilinositol-3-quinasa (PI3K) y la proteína quinasa B (Akt) que desempeña funciones críticas en los osteoblastos y formación ósea ya que regula procesos celulares fundamentales. La interacción entre E2/REα activa la vía de señalización PI3K-Akt donde la proteína PI3K es una enzima heterodimérica compuesta por una subunidad catalítica (P110) y una reguladora (p85), que son necesarias para realizar un amplio espectro de actividades celulares incluido el metabolismo y el envejecimiento. Por otro lado, Akt es una proteína quinasa de serina-treonina dependiente de fosfoinosítido. La interacción posterior de PI3K-Akt son reguladores cruciales de la reabsorción ósea por parte de los osteoclastos y de la formación ósea por parte de los osteoblastos, al promover su diferenciación y supervivencia para el mantenimiento y recambio de la masa ósea. La deficiencia de Akt en osteoblastos induce un fenotipo susceptible a apoptosis, además de suprimir la función y diferenciación celular, por lo tanto, la vía de señalización PI3K-Akt juega un papel clave en el proceso de formación de hueso en la membrana celular (44). Por otro lado, la interacción E2/RE promueve la activación de la vía de señalización de las proteínas quinasas activadas por mitógenos (MAPK) que son un conjunto de quinasas serina/treonina que regulan una amplia gama de estímulos. ERK presenta dos isoformas, ERK1 (MAPK3) y ERK2 (MAPK1), ambas expresadas en osteoblastos. ERK es activado por MAP2Ks-MEK1 (MAP2K1) y MEK2 (MAP2K2). Ratones con deleción de la línea germinal de Erk1 y una deleción condicional de Erk2 en el mesénquima de las extremidades (ratones Erk-1-/-Erk2Prx1), incluidos los osteoblastos, muestran una reducción sustancial de la mineralización del hueso, lo que demuestra la importancia de ERK para la mineralización de osteoblastos. Del mismo modo, los ratones que expresan la mutación de MEK1 dominante en osteoblastos muestran una masa ósea baja e hipomineralización de la clavícula y la bóveda craneal. En particular, estos ratones también muestran hipomineralización clavicular y craneal, los cuales son vestigios de ratones y humanos haploinsuficientes para Runx2, el regulador maestro de la diferenciación de osteoblastos (45) (Fig. 2). Otra vía de señalización estimulada por la unión de la testosterona con el receptor de andrógenos es el sistema renina angiotensina (RAS). Se ha reportado que RAS es un sistema complejo que funciona como mediador entre la formación y reabsorción del hueso a través de diversos mecanismos. El papel de RAS inicia con la conversión de angiotensinógeno en angiotensina I (AngI) la cual es activada por la renina, una proteasa altamente selectiva secretada por las células yuxtaglomerulares del riñón. Posteriormente, la AngI se convierte en angiotensina II (AngII) a través de la enzima convertidora de angiotensina (ECA). La relación entre el sistema renina angiotensina y el metabolismo óseo se basa principalmente en la regulación de la AngII en el hueso. Se ha reportado que AngII se asocia con un aumento significativo de los osteoclastos positivos para TRAP y con regulación positiva para de la expresión de RANKL a través de la quinasa extracelular de los osteoblastos (46) (Fig. 3). Sin embargo, estos efectos son reprimidos con tratamiento dirigido a la inhibición de la ECA o con bloqueadores de los receptores angiotensina tipo 1 (BRA) por lo que la vía de señalización RAS ha surgido como una estrategia en el tratamiento de trastornos del metabolismo óseo como la OP (47). Hoy en día el tratamiento de la OP en hombres no es diferente con respecto al indicado en mujeres y actualmente se han realizado pocos estudios sobre la eficacia de los fármacos en hombres. El tratamiento no farmacológico de la OP se basa esencialmente en estilos de vida y tampoco difiere entre hombres y mujeres (48). Sin embargo, la Endocrine Society ha formulado directrices específicas para el tratamiento de la OP masculina como los bifosfonatos, dirigidos a pacientes con fractura reciente de cadera y teriparatida para pacientes con problemas gastrointestinales y alto riesgo de fractura. Por otro lado, la North American Menopause Society (NAMS) ha sugerido que en el tratamiento de mujeres posmenopáusicas se implementen cambios en los hábitos alimenticios, estilo de vida e iniciar un tratamiento farmacológico con bifosfonatos como opciones de primera línea y raloxifeno en mujeres posmenopáusicas más jóvenes, para prevenir la pérdida de masa ósea y reducir el riesgo de fracturas vertebrales (49,50).
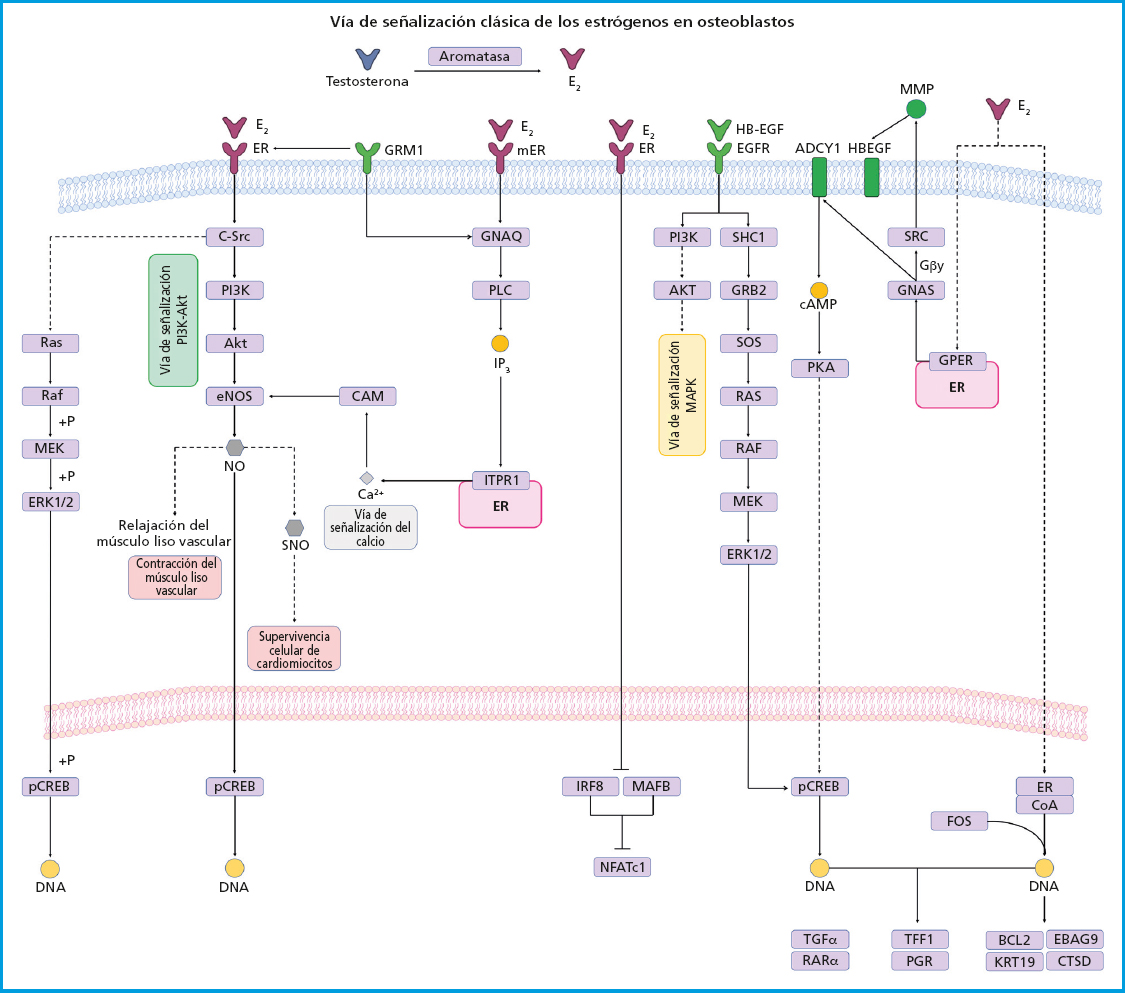
Figura 2. Señalización del receptor de estrógenos. Los estrógenos se difunden a través de la membrana plasmática y se une a REα o REβ que se dimerizan y translocan al núcleo. Los RE se unen a secuencias específicas, reclutan coactivadores y transcriben sus genes diana. Los RE unidos a estrógenos también pueden interactuar con factores de transcripción como AP1, SP1 y NF-kB, que también juegan un papel importante en la regulación de la osteoclastogénesis. La activación de los receptores de tirosina quinasa (EGFR) y los receptores acoplados a proteína G (GPCR) conducen a la activación de MAPK, PKA y PI3K-Akt.
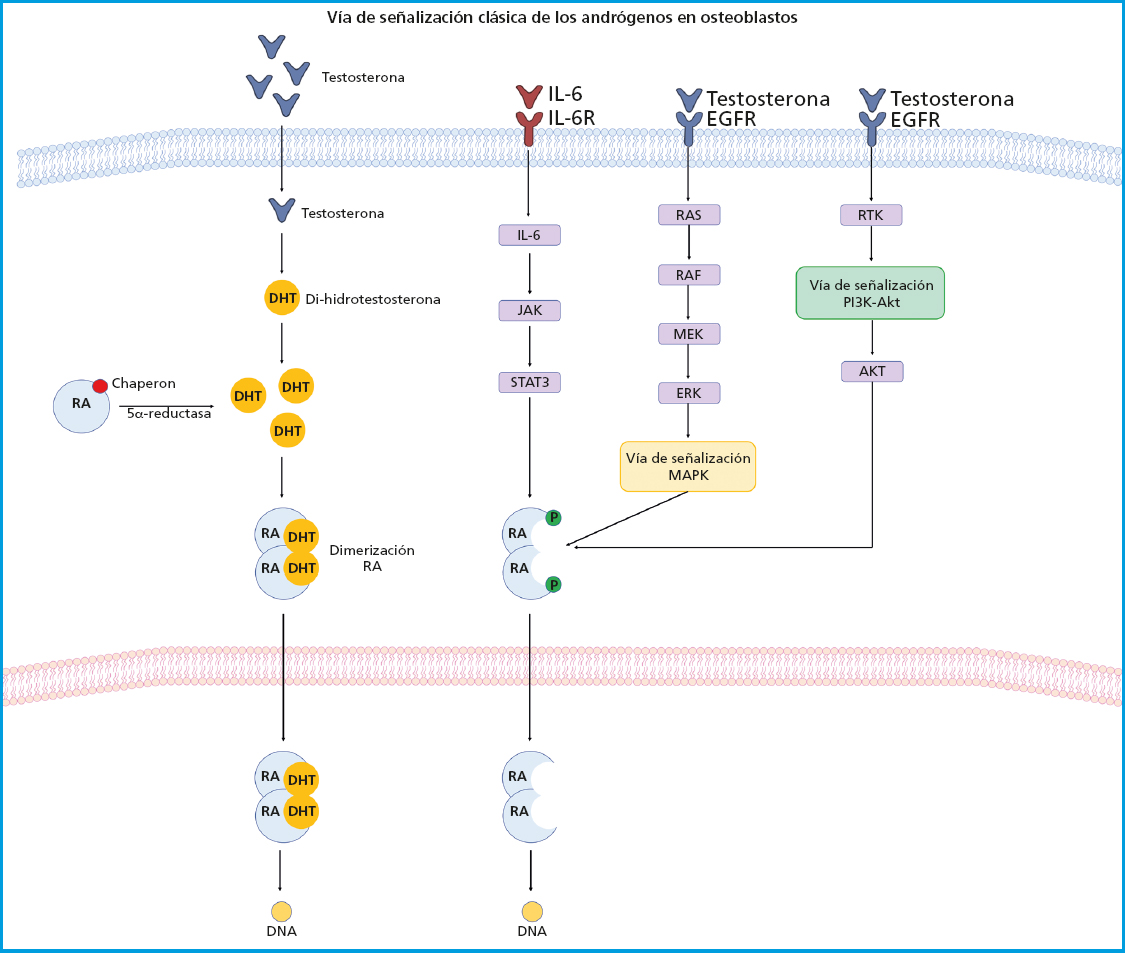
Figura 3. Señalización del receptor de andrógenos. El receptor de andrógenos (RA) se mantiene inactivo en el citoplasma por proteínas chaperonas. La unión de los andrógenos al RA conduce a la disociación del complejo chaperón lo que provoca un cambio conformacional en el RA seguido de su dimerización. El dímero del RA se traslada al núcleo y se une a promotores/potenciadores del gen diana facilitando a través de sus interacciones con los correguladores. Los RA también pueden afectar la señalización celular sin unirse directamente a los promotores de genes. En ausencia de andrógenos, varios factores de crecimiento y citocinas también pueden activar el AR al regular múltiples vías de señalización entre las que se encuentran la vía PI3K-Akt, MAPK/ERK mediada por RAS, las cuales están implicadas en la formación del hueso.
Por otro lado, el efecto de los andrógenos sobre marcadores subrogados como el TBS o el micro-CT han sido poco estudiado. En un trabajo realizado por Cauley y cols. en 2021 se analizó la DMO de 211 hombres mayores que recibieron tratamiento con testosterona moderadamente baja, sin otra razón que la edad. Se reportó que el tratamiento con testosterona durante un año, con respecto a un grupo control, aumentó notablemente los niveles de DMO volumétrica trabecular. Los resultados fueron analizados a través de una tomografía computarizada cuantitativa (QCT) en cadera y columna, observando un aumento en la resistencia ósea estimada. Sin embargo, los autores mencionan que las exploraciones QCT son costosas y presentan altos niveles de radiación, y es poco probable que se incorporen a la práctica clínica habitual, por lo que proponen el uso de la puntuación ósea trabecular (TBS) como medida indirecta de la microarquitectura ósea de columna vertebral, que se puede obtener a partir de análisis de textura de exploraciones rutinarias de DXA de columna lumbar y que, en conjunto con la herramienta de predicción FRAX®, pueden aumentar la precisión de la predicción de fracturas y mejorar el manejo clínico personalizado de la OP (51). Por otro lado, un estudio realizado por Movérare y cols. en 2003 buscó comparar el efecto de la activación de RE en el hueso in vivo con el efecto de activación de RA en ratones de 9 meses de edad orquiectomizados de tipo silvestre y con RE inactivado con el andrógeno 5α-dihidrotestosterona. Los análisis QCT de DMO demostraron que el efecto de conservación del hueso trabecular de la activación de RE y la activación de RAfue de la misma magnitud. Sin embargo, un análisis más detallado de la microarquitectura del hueso trabecular, con micro-CT de alta resolución, demostró que la activación de RE, pero no de RA, preservaba el espesor trabecular, mientras que la activación de RA solo preservaba el número de trabéculas (52). Por lo tanto, el uso de estas herramientas puede ser empleada en para crear simulaciones por computadora del remodelado óseo y evaluar dinámicamente una respuesta a la terapia con testosterona en la práctica clínica habitual.
CONCLUSIÓN
El hueso es un tejido que se encuentra en constante renovación gracias al proceso de reabsorción y formación ósea. Sin embargo, alteraciones en este proceso pueden conducir al desarrollo de enfermedades como la OP. Aunque muchos estudios han reconocido a los estrógenos y su interacción con receptores específicos como reguladores del metabolismo óseo, los andrógenos han sido poco explorados. La evidencia sugiere que andrógenos como la testosterona juegan un papel importante en el mantenimiento de la DMO y la salud ósea en hombres. Además, se ha identificado que muchos mecanismos moleculares de la testosterona actúan sobre vías de señalización implicadas en el metabolismo óseo entre las que se encuentran la vía, PI3K-Akt, MAPK y RAS, las cuales han sido previamente descritas por su papel en el mantenimiento de la masa ósea. Por lo que el papel de la testosterona podría explorarse como una opción de tratamiento para mejorar la DMO en hombres mayores.

