
ORIGINAL
Estudio funcional de los polimorfismos del promotor del gen de la esclerostina
Functional study of promoter gene polymorphisms of sclerostin
Pérez-Campo F.M.2, Sañudo C.1, Krebesova R.1, Delgado-Calle J.3 y Riancho J.A.1
1 Departamento de Medicina Interna - Hospital U.M. Valdecilla - Universidad de Cantabria - IDIVAL - Santander (España)
2 Departamento de Biología Molecular - Universidad de Cantabria - Santander (España) ]]>
3 Departamento de Anatomía y Biología Celular - Facultad de Medicina de la Universidad de Indiana - Centro Médico de Administración de Veteranos Roudebush - Indianápolis - Indiana (EE.UU.)
Este trabajo se realizó gracias a una beca de la FEIOMM 2013 "Interacción entre Osterix, Runx2 y Esclerostina: mecanismos moleculares y repercusión sobre la masa ósea" y del Instituto de Salud Carlos III (PI12-615), la cual puede ser cofinanciada con fondos FEDER de la Unión Europea.
Trabajo premiado con una beca de Investigación de Biología Molecular Ósea FEIOMM 2013.
Dirección para correspondencia
RESUMEN
La esclerostina, codificada por el gen SOST, es un inhibidor de la vía Wnt, y por ello tiene una influencia negativa sobre la masa ósea. Algunos polimorfismos del promotor de SOST se han asociado a diferencias en la densidad mineral ósea (DMO) en varios estudios, pero se desconocen cuáles son los mecanismos moleculares implicados. El objetivo de este estudio fue comprobar las consecuencias funcionales de uno de esos polimorfismos in vitro. Para ello se clonó la región promotora proximal del gen SOST, con diferentes alelos del polimorfismo rs851054, y se transfectaron en células HEK-293T, SAOS-2 y HOS-TE85. En ningún caso se observaron diferencias significativas en la actividad transcripcional entre los vectores con el alelo A y los vectores con el alelo G. La co-transfección de vectores de expresión de los factores de transcripción RUNX2 y OSX estimuló claramente la actividad transcripcional (2,5±0,9 veces sobre el valor basal para el alelo A y 1,9±0,8 veces para el alelo G; en ambos casos, p<0,05), sin que hubiera, sin embargo, diferencias entre los alelos. Tampoco se hallaron diferencias en la fijación a proteínas nucleares analizadas en experimentos de retardo de la movilidad electroforética.
En conclusión, la región situada antes del inicio de la traducción del gen SOST tiene una potente actividad promotora, que es aumentada por los factores RUNX2 y OSX. Las variantes frecuentes de esta región se han asociado con la DMO, pero los mecanismos implicados son aún desconocidos, puesto que los alelos analizados no muestran diferencias en la actividad transcripcional in vitro.
Palabras clave: esclerostina, regulación génica, polimorfismos, transfección.
Sclerostin, encoded by the SOST gene, inhibits the Wnt pathway and, consequently, tends to decrease bone mass. Some polymorphisms of the SOST promoter have been associated with bone mineral density (BMD), but the molecular mechanisms involved are unknown. The aim of this study was to study the functional role of one polymorphism in vitro. We cloned the proximal promoter region of SOST gene, containing different alleles at the rs851054 SNP, in luciferase reporter vectors and transfected them into the cell lines HEK-293T, SAOS-2 and HOS-TE85. We did not find significant differences in the transcriptional activity of vectors with either the A or the G allele of the SNP. The co-transfection of vectors expressing RUNX2 and OSX markedly increased the transcriptional activity of the SOST promoter constructs (A allele, 2.5±0.9 fold, p<0.05; G allele, 1.9±0.8 fold, p<0.05), without significant differences between the rs851054 alleles. Moreover, no allele differences were detected in EMSAs.
In conclusion, the DNA region upstream of the TSS of the SOST gene has a strong promoter activity that is enhanced by RUNX2 and OSX. Frequent allelic variants in this region have been associated with BMD, but the mechanisms involved remain to be elucidated because no functional differences between alleles were detected in vitro.
Key words: sclerostin, gene regulation, polymorphisms, transfection.
Introducción
La evolución de la masa ósea a largo plazo viene determinada por el balance entre la resorción y la formación de hueso, de manera que una actividad formadora de hueso insuficiente para sustituir el destruido durante la resorción inevitablemente conduce a la disminución del tejido óseo, característica propia de la osteoporosis. Los osteoblastos, encargados de la formación ósea, derivan de células troncales mesenquimales que pueden dar lugar también a otros tipos celulares, como adipocitos o condrocitos. La proliferación y la diferenciación de los precursores osteoblásticos está controlada por diferentes factores, intracrinos, paracrinos y endocrinos. Así, algunos factores de transcripción, como RUNX2 y Osterix (OSX), se consideran esenciales en las primeras etapas de la diferenciación osteoblástica. La vía Wnt desempeña también un papel importante1. Los ligandos Wnt se unen a sus receptores presentes en la membrana celular y, a través de diversos mediadores intracelulares, modifican la expresión de sus genes diana que, en general, actúan favoreciendo la formación ósea y aumentando la expresión de OPG, lo que secundariamente resulta en una disminución de la resorción. Los receptores de los ligandos Wnt son complejos moleculares que incluyen al menos dos proteínas: Frizzled y LRP, de las cuales existen varias formas2,3. Como en el caso de muchos otros sistemas reguladores, también existen inhibidores de la vía Wnt. Uno de los más estudiados es la esclerostina, codificada por el gen SOST. Este gen se expresa de manera preferente en los osteocitos4-8. Siendo un inhibidor de la vía Wnt, la esclerostina ejerce una influencia negativa sobre la masa ósea. Su papel en la biología esquelética parece importante. De hecho, los anticuerpos neutralizantes de la esclerostina ejercen un potente efecto anabólico sobre el esqueleto, y las mutaciones inactivadoras del gen SOST provocan un aumento exagerado de la masa ósea9-11.
En consonancia con el papel biológico de la esclerostina, las variantes alélicas del gen SOST parecen influir en la masa ósea. Así, en algunos estudios de barrido genómico (GWAS) y en otros estudios de asociación genética, se ha encontrado una relación entre algunos polimorfismos frecuentes del gen SOST y la densidad mineral ósea (DMO). De hecho, nuestro grupo encontró una asociación entre unos polimorfismos situados en la región promotora de SOST y la DMO en mujeres postmenopáusicas12. El objetivo del presente estudio fue explorar la capacidad reguladora de uno de esos polimorfismos mediante estudios funcionales in vitro. Para ello hemos utilizado vectores en los que el promotor de SOST se ha insertado delante de un gen reportero que codifica una proteína (luciferasa) cuya actividad es fácilmente medible, y hemos realizado experimentos para comprobar la capacidad de fijación de proteínas nucleares a esas regiones valorando el retardo de la movilidad electroforética.
Material y métodos
]]> Construcción de vectores reporterosEl promotor de SOST (1-1440) se clonó a partir de ADN genómico de dos individuos con genotipo conocido, homocigotos para los alelos A y G, respectivamente, del polimorfismo rs851054 (Figura 1). Ambos fragmentos tenían, por lo demás, secuencias idénticas e iguales a la secuencia de referencia del genoma humano. La extracción de ADN se produjo tras el consentimiento informado, en el seno de un estudio de factores genéticos implicados en la osteoporosis, autorizado por el Comité de Ética en Investigación Clínica. Para la amplificación de los correspondientes fragmentos genómicos por PCR se utilizaron cebadores diseñados con el programa Primer3 que incluían secuencias para las enzimas de restricción Smal y Xhol, a fin de facilitar su posterior clonaje. Se cortaron con estas enzimas los amplicones y el vector PGL2 y se unieron los fragmentos mediante T4 DNA Ligasa (NE Biolabs). Los vectores se transfectaron en células competentes DH5α (Invitrogen), las cuales se cultivaron después en placas con agar y medio suplementado con ampicilina. Las colonias seleccionadas se expandieron en cultivo líquido, y posteriormente se extrajeron los plásmidos y se comprobó que contenían los insertos correctos, sin mutaciones, mediante secuenciación convencional. Por tanto, en estos vectores se encontraba la secuencia promotora de SOST dirigiendo la transcripción del gen reportero, codificador de luciferasa. Así, la medida de la actividad luciferasa era reflejo de la actividad promotora de la transcripción de la secuencia del alelo del promotor SOST que se clonó en el vector.
Transfección y análisis de la actividad transcripcional
Las células HEK-293T (una línea derivada de riñón humano) y las líneas osteoblásticas humanas SaOS-2 y HOS-TE85 se transfectaron con los vectores conteniendo las secuencias promotoras de SOST y un vector control RSV-βGAL, que expresa constitutivamente el gen LacZ, que codifica para la β-galactosidasa, a fin de normalizar los resultados en función de la eficiencia de la transfección. Como control negativo y en paralelo se realizó también la transfección con el vector vacío. Para la transfección se sembraron 125.000 células HEK-293T (o 50.000 SAOS-2 o HOS-TE85) en cada pocillo de una placa de 24 pocillos. Una vez alcanzada una confluencia del 80% se transfectaron 500 ng de los vectores, utilizando Lipofectamina 3000, según las recomendaciones del fabricante (Invitrogen). A las 48 horas se aspiró el medio y se lisaron las células con 70 µl de tampón, tras lo que se midió la actividad de galactosidasa (Galacto-Light PlusTM β-Galactosidase Reporter Gene Assay System, Applied Biosystems) y de luciferasa (Luciferase Assay System, Promega) mediante luminometría. La co-transfección con vectores de expresión de RUNX2 y de OSX13 se realizó siguiendo un procedimiento similar, pero asegurando que la cantidad total de ADN exógeno a transfectar se mantenía constante en todos los pocillos.
De cada transfección se realizaron duplicados y triplicados técnicos. Además cada experimento se repitió en al menos tres ocasiones diferentes para obtener triplicados biológicos. Los resultados se expresaron como el cociente entre la actividad luciferasa y la actividad galactosidasa en los lisados celulares.
Análisis de la fijación de proteínas nucleares
Los extractos proteicos se obtuvieron a partir de 50x106 células HOS-TE85. Para ello, se lisaron en un tampón conteniendo inhibidores de proteasas (50 mM KCl, 0,5% NP-40, 25 mM HEPES, 1,5 pM Leupeptina, 46,88 µM Aprotinina, 125 µM DTT, 1 mM PMSF), y se centrifugaron durante 1 minuto a 10.000 rpm a 4oC. Tras lavarlo, se resuspendieron las células en 100 µl de tampón de extracción (500 mM KCl, 25 mM HEPES, 10% glicerol, 1,5 pM Leupeptina, 46,88 µM Aprotinina, 125 µM DTT, 1 mM PMSF) y se centrifugó durante 5 minutos a 14.000 rpm a 4oC. Se recuperó el sobrenadante, se cuantificó la concentración de proteínas y se ajustó a 5 µg/µl.
]]> Para el ensayo de retardo de la movilidad electroforética (EMSA) se utilizaron parejas de oligonucleótidos, incluyendo marcaje con IR dye 700 en el extremo 5' del componente "forward" de cada pareja (Biolegio). La secuencias fueron las siguientes:rs851054 alelo A Fwd: AACAGAAACACCTTGGGCCA
rs851054 alelo A Rev: TGGCCCAAGGCGTTTCTGTT
rs851054 alelo G Fwd: AACAGAAACGCCTTGGGCCA
rs851054 alelo G Rev: TGGCCCAAGGCGTTTCTGTT
Para el retardo se utilizaron geles prefabricados de poliacrilamida al 6% (Invitrogen) y el kit Odissey Infrared EMSA. Tras anillar los oligonucleótidos se incubaron con el extracto proteico durante 20 minutos, se cargaron en el gel y se corrieron a 70 V durante 60 minutos, tras lo que se capturaron las imágenes de los geles. En estos experimentos, si las sondas fijan proteínas nucleares se produce un retardo en su movilidad electroforética en comparación con la migración experimentada por la sonda aislada.
Resultados
Las construcciones con la secuencia promotora de SOST mostraron una elevada capacidad activadora de la transcripción, aumentando los niveles de expresión de luciferasa hasta 1.000 veces en comparación con los vectores vacíos. La actividad fue aparentemente mayor en las células HEK-293T que en las otras líneas (Figura 2).
]]>

Sin embargo, no encontramos diferencias significativas entre los dos alelos del promotor de SOST en ninguna de las líneas celulares analizadas. De hecho, la relación entre la actividad transcripcional de los alelos A y G (cocientes A/G) en las células SAOS-2, HOS-TE85 y 293T fueron de 1,3±0,7, 1,0±0,4 y 0,8±0,7, respectivamente. En ninguno de los tres resultó significativamente diferente de la unidad (Figura 2).
La co-transfección de vectores que expresan constitutivamente RUNX2 y OSX aumentó la actividad transcripcional del promotor de SOST (en promedio, 2,5±0,9 veces sobre el valor basal para el alelo A y 1,9±0,8 veces para el alelo G; en ambos casos p<0,05). Este aumento también resultó ser independiente de qué alelo estaba presente en el promotor de SOST (Figura 3).
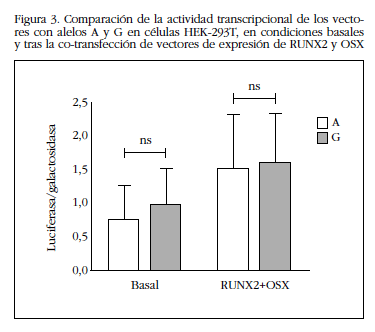
Por otro lado, en los experimentos de retardo de la movilidad electroforética comprobamos que la región donde radica ese polimorfismo era capaz de fijar proteínas nucleares, presumiblemente con función reguladora, sin que observáramos diferencias entre ambos alelos (Figura 4). Por ello, no se efectuaron estudios ulteriores para identificar la naturaleza de esas proteínas.

Discusión
El papel de la esclerostina en la biología ósea es indudable, como lo revelan el marcado aumento de la DMO que se observa tras la administración de romosozumab, un anticuerpo monoclonal que bloquea la acción de la esclerostina, y la masa ósea excesiva que presentan los pacientes con esclerosteosis o con enfermedad de Van Buchem, portadores de mutaciones poco frecuentes que inducen una pérdida de función del gen SOST, poco frecuentes14. Aunque la evidencia es menor, varios estudios sugieren que las variantes alélicas frecuentes del gen pueden influir también, en menor grado, sobre la DMO y el riesgo de sufrir fracturas osteoporóticas. De hecho, en un estudio de barrido genómico con un elevado tamaño muestral se encontró una asociación del polimorfismo rs4792909 con la DMO15. Este polimorfismo se encuentra en una región intergénica, siendo SOST el gen más cercano, situado a unas 40 kb. Por otro lado, varios grupos, incluido el nuestro, han analizado la relación de algunos polimorfismos frecuentes de la región promotora de SOST con la DMO. Así, nosotros hallamos una asociación significativa de los polimorfismos rs851054 y rs851056, separados sólo por 560 pb e integrantes de un mismo bloque haplotípico con la DMO de columna en mujeres12. Algunos autores han confirmado la asociación de los polimorfismos del promotor de SOST con la DMO en otras poblaciones16-20. Por otro lado, en un estudio reciente que incluyó un pequeño grupo de pacientes, se encontró una asociación de los alelos de rs851054 con la expresión de SOST en el tejido óseo21.
El objetivo principal de este estudio fue explorar la posible repercusión funcional de uno de esos polimorfismos, en concreto se trató de analizar su influencia sobre la actividad transcripcional del promotor del gen SOST. Los experimentos de transfección con vectores reporteros y los de retardo de la movilidad electroforética no han revelado diferencias entre las variantes alélicas estudiadas. Son diversas las limitaciones de los modelos utilizados y por ello hay varios motivos que podrían explicar estos resultados negativos. En primer lugar, podría plantearse que el modelo in vitro no refleja adecuadamente la situación in vivo. Por ejemplo, no puede excluirse la posibilidad de que la actividad reguladora requiera de regiones genómicas con actividad enhancer, alejadas del promotor y no incluidas, por tanto, en la región clonada en los vectores. Por otro lado, al menos teóricamente podría pensarse que la diferencia en la actividad de los alelos sólo se manifiesta en respuesta a factores reguladores que únicamente se expresan en determinados tipos celulares, como los osteocitos, mientras que están ausentes en los tipos celulares utilizados en nuestros experimentos. En segundo lugar, cabe pensar que la asociación con la DMO pudiese depender de otros polimorfismos frecuentes, en desequilibrio de unión con los aquí estudiados. De ser así, deberían encontrarse en regiones lejanas, alejadas del promotor, puesto que en esta región existe un fuerte ligamiento. En este sentido, cabe señalar que hay una región reguladora de SOST situada a unas 50 kb, que contiene algunos polimorfismos que se han relacionado de manera inconsistente con la DMO20,22. Esa región es capaz de fijar el factor de transcripción MEF2C y desempeña un papel importante en la regulación de SOST, porque su falta ocasiona el fenotipo de la enfermedad de Van Buchem en humanos y un incremento de la masa ósea en modelos murinos23,24. Una tercera posibilidad es que la asociación con la DMO dependa de algunos polimorfismos de baja frecuencia, situados en la región promotora. Finalmente, los polimorfismos podrían actuar a través de mediadores epigenéticos que no pueden ser recapitulados adecuadamente en los modelos in vitro. Por tanto, se precisan otros estudios, incluyendo el análisis sistemático de las variantes de las regiones 5' y 3' del gen y de otras que se encuentren asociadas a él teniendo en cuenta la estructura tridimensional de la cromatina, así como estudios de mutagénesis en vivo para esclarecer los mecanismos por los que estas variantes alélicas pueden modular la actividad de la esclerostina y asociarse a diferencias en la masa ósea25.
En este estudio hemos corroborado los resultados previos que indican que RUNX2 y OSX aumentan la expresión de SOST en humanos13. Eso indica que estos factores de transcripción tienen un papel complejo sobre la línea osteoblástica. Por un lado, está bien establecido su papel como determinantes de las etapas iniciales de diferenciación de las células mesenquimales hacia osteoblastos, lo que posibilita que haya un número adecuado de osteoblastos formadores de hueso. Por otro, promueven la actividad del promotor de SOST. Por tanto, en células capaces de expresar este gen, como los osteocitos, podrían promover la secreción de esclerostina y así contribuir a evitar una formación ósea exagerada. En todo caso, las variantes del promotor de SOST no parecen influir en la respuesta a estos factores.
En conclusión, en este estudio hemos confirmado que la región situada antes del inicio de la traducción del gen SOST tiene una potente actividad promotora, que es además inducida por los factores de transcripción RUNX2 y OSX. Las variantes frecuentes de esta región se han asociado con la masa ósea, pero los mecanismos implicados son aún desconocidos, puesto que los alelos no muestran diferencias en la actividad transcripcional in vitro.
Agradecimientos
Los autores agradecen al Dr. Svante Paabo el generoso suministro del plásmido de expresión de RUNX2.
]]> Declaración de intereses:
Los autores declaran no tener conflictos de intereses.
Bibliografía
1. Riancho JA, Hernandez JL. Pharmacogenomics of osteoporosis: a pathway approach. Pharmacogenomics. 2012;13(7):815-29. [ Links ]
2. Baron R, Rawadi G. Wnt signaling and the regulation of bone mass. Curr Osteoporos Rep. 2007;5:73-80. [ Links ]
3. Bodine PV, Komm BS. Wnt signaling and osteoblastogenesis. Rev Endocr Metab Disord. 2006;7:33-9. [ Links ]
]]>4. Semenov M, Tamai K, He X. SOST is a ligand for LRP5/LRP6 and a Wnt signaling inhibitor. J Biol Chem. 2005;280(29):26770-5. [ Links ]
5. Li X, Zhang Y, Kang H, Liu W, Liu P, Zhang J, et al. Sclerostin binds to LRP5/6 and antagonizes canonical Wnt signaling. J Biol Chem. 2005;280:19883-7. [ Links ]
6. Moester MJ, Papapoulos SE, Lowik CW, van Bezooijen RL. Sclerostin: current knowledge and future perspectives. Calcif Tissue Int. 2010;87:99-107. [ Links ]
7. Ott SM. Sclerostin and Wnt signaling--the pathway to bone strength. J Clin Endocrinol Metab. 2005;90:6741-3. [ Links ]
8. Poole KE, van Bezooijen RL, Loveridge N, Hamersma H, Papapoulos SE, Lowik CW, et al. Sclerostin is a delayed secreted product of osteocytes that inhibits bone formation. FASEB J. 2005;19:1842-4. [ Links ]
]]>9. Loots GG, Kneissel M, Keller H, Baptist M, Chang J, Collette NM, et al. Genomic deletion of a long-range bone enhancer misregulates sclerostin in Van Buchem disease. Genome Res. 2005;15:928-35. [ Links ]
10. Balemans W, Van Hul W. Identification of the disease-causing gene in sclerosteosis--discovery of a novel bone anabolic target? J Musculoskelet Neuronal Interact. 2004;4:139-42. [ Links ]
11. Kim CA, Honjo R, Bertola D, Albano L, Oliveira L, Jales S, et al. A known SOST gene mutation causes sclerosteosis in a familial and an isolated case from Brazilian origin. Genet Test. 2008;12:475-9. [ Links ]
12. Valero C, Zarrabeitia MT, Hernandez JL, Pineda B, Cano A, Garcia-Perez MA, et al. Relationship of sclerostin and secreted frizzled protein polymorphisms with bone mineral density: an association study with replication in postmenopausal women. Menopause. 2011;18:802-7. [ Links ]
13. Perez-Campo FM, Santurtun A, Garcia-Ibarbia C, Pascual MA, Valero C, Garces C, et al. Osterix and RUNX2 are Transcriptional Regulators of Sclerostin in Human Bone. Calcif Tissue Int. 2016;99(3):302-9. [ Links ]
]]>14. Balemans W, Van Hul W. Human genetics of SOST. J Musculoskelet Neuronal Interact. 2006;6:355-6. [ Links ]
15. Estrada K, Styrkarsdottir U, Evangelou E, Hsu YH, Duncan EL, Ntzani EE, et al. Genome-wide meta-analysis identifies 56 bone mineral density loci and reveals 14 loci associated with risk of fracture. Nat Genet. 2012;44:491-501. [ Links ]
16. Kuipers AL, Zhang Y, Yu S, Kammerer CM, Nestlerode CS, Chu Y, et al. Relative influence of heritability, environment and genetics on serum sclerostin. Osteoporos Int. 2014;25(3):905-12. [ Links ]
17. Sims AM, Shephard N, Carter K, Doan T, Dowling A, Duncan EL, et al. Genetic analyses in a sample of individuals with high or low bone density demonstrates association with multiple Wnt pathway genes. J Bone Miner Res. 2008;23:499-506. [ Links ]
18. Yerges LM, Klei L, Cauley JA, Roeder K, Kammerer CM, Moffett SP, et al. High-density association study of 383 candidate genes for volumetric BMD at the femoral neck and lumbar spine among older men. J Bone Miner Res. 2009;24:2039-49. [ Links ]
]]>19. Mencej-Bedrac S, Prezelj J, Kocjan T, Komadina R, Marc J. Analysis of association of LRP5, LRP6, SOST, DKK1, and CTNNB1 genes with bone mineral density in a Slovenian population. Calcif Tissue Int. 2009;85:501-6. [ Links ]
20. Uitterlinden AG, Arp PP, Paeper BW, Charmley P, Proll S, Rivadeneira F, et al. Polymorphisms in the sclerosteosis/van Buchem disease gene (SOST) region are associated with bone-mineral density in elderly whites. Am J Hum Genet. 2004;75:1032-45. [ Links ]
21. Lhaneche L, Hald JD, Domingues A, Hannouche D, Delepine M, Zelenika D, et al. Variations of SOST mRNA expression in human bone are associated with DNA polymorphism and DNA methylation in the SOST gene. Bone. 2016;92:107-15. [ Links ]
22. Balemans W, Foernzler D, Parsons C, Ebeling M, Thompson A, Reid DM, et al. Lack of association between the SOST gene and bone mineral density in perimenopausal women: analysis of five polymorphisms. Bone. 2002;31:515-9. [ Links ]
23. Collette NM, Genetos DC, Economides AN, Xie L, Shahnazari M, Yao W, et al. Targeted deletion of Sost distal enhancer increases bone formation and bone mass. Proc Natl Acad Sci. U S A 2012;109(35):14092-7. [ Links ]
]]>24. Balemans W, Cleiren E, Siebers U, Horst J, Van Hul W. A generalized skeletal hyperostosis in two siblings caused by a novel mutation in the SOST gene. Bone. 2005;36:943-7. [ Links ]
25. Huang Q. Genetic study of complex diseases in the post-GWAS era. J Genet Genomics. 2015;42(3):87-98. [ Links ]
![]() Dirección para correspondencia:
Dirección para correspondencia:
José A. Riancho
Departamento de Medicina Interna
Hospital U.M. Valdecilla ]]>
Avda. Valdecilla, sn
39008 Santander (España)
Correo electrónico: rianchoj@unican.es
Fecha de recepción: 16/07/2016
Fecha de aceptación: 25/09/2016