










My SciELO
 Custom services
Custom servicesServices on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Española de Enfermedades Digestivas
Print version ISSN 1130-0108
Rev. esp. enferm. dig. vol.97 n.9 Madrid Sep. 2005
| TRABAJOS ORIGINALES |
Modificaciones inmunopatológicas de la mucosa rectal en un modelo animal de alergia alimentaria
M. Vinuesa, N. Bassan, S. Roma y F. Pérez
Cátedra de Histología y Embriología. Facultad de Ciencias Médicas. Universidad Nacional de Rosario. Rosario, Santa Fe. Argentina
RESUMEN
Objetivo: determinar las modificaciones inmunopatológicas en la mucosa rectal de conejo sensibilizado con ovoalbúmina (OVA) y desafiado localmente.
Diseño experimental: treinta conejos divididos en tres grupos G1: normal; G2 sensibilizado por vía subcutánea con OVA y G3: sensibilizado y desafiado localmente con OVA y muestreados 4 horas después del desafío. Los niveles de IgE anti-OVA específica fueron evaluados por el test de anafilaxia cutánea pasiva (PCA). Se contaron 200 campos de mayor aumento en cada grupo. Los resultados fueron expresados como media aritmética y error standard aplicándose el test de la t de
Student.
Resultados: CD 4: G1: 8,3 ± 0,06; G2: 13,4 ± 0,08 y G3: 8,25 ± 0,06. CD 5: G1: 7,3 ± 0,05; G2: 9,4 ± 0,05 y G3: 11,3 ± 0,06. CD 25: G1: 13., ± 0,08; G2: 15,1 ± 0,13 y G3: 25,5 ± 0,15. Cadena
µ: G1: 10,4 ± 0,06; G2: 3,8 ± 0,02 y G3: 6,0 ± 0,10. RLA II (DR): G1: 11,6 ± 0,O5; G2: 19,2 ± 0,09 y G3: 19,1 ± 0,11. En todos los casos los grupos G2 y G3 presentaron diferencias estadísticamente significativas
versus G1 (p < 0,001).
Conclusión: observamos un incremento en el número de células CD25+ (receptor interleuquina-2) en G3 y una disminución en células cadena
µ positivas en G2 y G3, probablemente debido a la activación de células B y expresión de otras inmunoglobulinas en superficie celular. La expresión de RLA II fue mayor en G2 y G3. Este receptor es considerado un marcador de activación expresado por macrófagos, células T y B. Concluimos que los datos obtenidos constituyen un aporte al conocimiento de la inmunopatología de la reacción anafiláctica local en mucosa rectal en animales sensibilizados por vía sistémica.
Palabras clave: Inmunopatología. Recto. Conejo. Alergia alimentaria.
INTRODUCCIÓN
Se denomina alergia alimentaria a respuestas inmunes anómalas que se desarrollan en individuos previamente sensibilizados, al incorporar por vía digestiva sustancias que contienen el alérgeno al cual están sensibilizados (1,2).
En el tubo digestivo los antígenos alimentarios pueden ser incorporados mediante una entrada apropiada de moléculas y producir una respuesta de tolerancia (3) o pueden ingresar en forma inapropiada (barrera mucosa alterada), produciendo sensibilización con altos niveles de IgE específica que se une a mastocitos y basófilos (4-8).
Un nuevo contacto del antígeno, a nivel del intestino en el individuo sensibilizado, desencadena una cascada de hechos que conducen a la generación de una reacción inflamatoria aguda anafiláctica local (9-11). Esta reacción presenta modificaciones cualitativas de la mucosa y cuantitativas de las células participantes en la referida respuesta inmune.
Se estima que el 0,2% de la población presenta síntomas de alergia a los alimentos. En EE.UU. el 5% de los menores de 3 años y el 1,5% de la población general (alrededor de 4.000.000) manifiesta síntomas de alergia a los alimentos (12).
Los alérgenos pueden ser componentes normales de los nutrientes, formar parte de los aditivos o no encontrarse en el producto natural, pero sí en variedades transgénicas de los mismos. El modo de preparación puede aumentar o disminuir la capacidad antigénica de un alimento o modificar estructuras moleculares dando lugar a reacciones cruzadas (13).
Los alimentos más estudiados, capaces de producir alergia alimentaria son: leche de vaca, huevo de gallina, trigo, tomate, chocolate, maní, kiwi, pescados, ostras y mariscos (14).
En las personas sensibilizadas al látex (Hevea Brasiliensis) se observa alergia cruzada con frutas, como banana y kiwi, que comparten proteínas o antígenos comunes con el látex (15,16). En este caso de sensibilización, por el uso de guantes para tacto rectal, sondas o preservativos en el coito anal, el recto pasa a ser un órgano de ingreso del antígeno, por vía digestiva. El conejo resulta un modelo biológico muy útil para el estudio de alergia alimentaria basado en distintas razones. Es un lagomorfo que presenta una importante conservación génica con los humanos (17). Produce altos niveles de IgE ante la sensibilización subcutánea (18-20). Además, realiza la cecotrofia en su fisiología digestiva, posibilitando que una importante cantidad de macromoléculas alimentarias potencialmente antigénicas alcancen sectores distales del intestino grueso (21).
El recto pertenece al sistema inmune mucoso común (SIMC), lo cual determina que los procesos de sensibilización y desafío posterior con antígenos alimentarios, generen modificaciones histopatológicas e inmunológicas capaces de ser evaluadas en su mucosa (22-24).
El objetivo del presente trabajo es establecer las modificaciones cuantitativas en poblaciones celulares participantes en la respuesta inmune a nivel de la mucosa rectal, en un modelo animal de alergia alimentaria.
MATERIALES Y MÉTODOS
Se estudiaron 30 conejos neocelandeces divididos en tres grupos: grupo 1: (n = 10) control normal; grupo 2: (n = 10) sensibilizado con ovoalbúmina (OVA) por vía subcutánea (SC); grupo 3 (n = 10) sensibilizado por vía subcutánea con OVA y desafiado por vía rectal con OVA.
Los conejos de los grupos 2 y 3 fueron sensibilizados por vía subcutánea en dos oportunidades, con un intervalo de 15 días, con 2 ml de una suspensión de 70 µg de OVA en 30 mg de ALUM/ml.
Quince días después de la última sensibilización, previo ayuno de 24 horas, los animales del grupo 3 fueron desafiados por vía rectal con una solución de 50 mg OVA en 5 ml de solución salina (21).
Los animales se sacrificaron por sobredosis de éter sulfúrico, 4 h después del desafío rectal, de acuerdo a las consideraciones aprobadas por el Comité de Ética de la Facultad de Ciencias Médicas y se obtuvieron las muestras de recto requeridas para las distintas técnicas.
Las muestras para inmunohistoquímica se sumergieron en medio líquido protector para congelación rápida (OCT) y se congelaron en nitrógeno líquido, se almacenaron a -20 ºC y se cortaron a 8 µg con crióstato. El material para histología convencional se incluyó en parafina y cortó a 6 µg.
Los anticuerpos monoclonales utilizados fueron:
-Ratón anti-conejo CD5 (KEN-5, BALB/c IgG 1, Biosource, Camarillo, California, EE.UU.), para la identificación de células T totales maduras y determinadas células B (25).
-Ratón anti-conejo CD4 (KEN-4, BALB/c IgG 2a, Biosource, Camarillo, California, EE.UU.) para la identificación de subpoblación de linfocitos T auxiliares CD4 positivos (25).
-Ratón anti-conejo CD25 (KEI-alfa 1, BALB/c IgG 2b, Pharmingen, San Diego, California, EE.UU.), receptor de la interleuquina-2, marcador de activación celular (25).
-Ratón anti-conejo inmunoglobulina M (cadena µ, NRBM BALB/c IgG 1, Biosource, Camarillo, California, EE.UU.) para la identificación de linfocitos B.
-Ratón anti-conejo RLA-DR (RDR34, BALB/c IgG 2b, Pharmingen, San Diego, California, EE.UU.) para la identificación de RLA DR (homólogo del HLA DR humano) marcador de activación en células presentadoras de antígeno.
Para la inmunotinción de las células positivas de utilizó el sistema de detección biotina-streptavidina (Serotec Ltd. Oxford, Reino Unido) y AEC como cromógeno.
Los cortes por congelación, se fijaron en acetona a 4 grados centígrados durante 10 minutos, se bloquearon con suero no inmune al 2% en solución tampón TRIS salina (TBS) para evitar tinciones inespecíficas y se incubaron durante 60 minutos a 37 ºC con los anticuerpos monoclonales mencionados.
Las células positivas fueron analizadas en 200 campos de 400x en cada grupo y expresados como la media aritmética por campo microscópico. Los resultados fueron analizados estadísticamente a través de la obtención de la media aritmética, error estándar y test de comparación de medias.
Se evaluaron los títulos de IgE específica sérica anti OVA de los conejos sensibilizados mediante el test de anafilaxia cutánea pasiva (PCA) (26).
RESULTADOS
La prueba de anafilaxia cutánea pasiva (PCA) demostró niveles detectables de IgE anti ovoalbúmina (IgE anti-OVA) en los grupos sensibilizados (G2 y G3) a una dilución de 1/160.
Desde el punto de vista histopatológico, en el grupo experimental sensibilizado y desafiado vía rectal con ovoalbúmina, se observó edema mucoso parcheado, imágenes de linfangiectasias e infiltración de eosinófilos. Estos últimos se distribuyeron en la mucosa subyacente al epitelio. En los otros grupos (G1) control y (G2) sensibilizados sin desafiar, no se presentaron modificaciones histopatológicas visualizables.
Los resultados de la cuantificación de las poblaciones celulares inmunes se presentan en la tabla I. Las células CD4 positivas (Fig. 1) predominan en la lámina propia del recto tanto de animales control como de los grupos experimentales, no observándose diferencias significativas entre el grupo que fue sometido a la reacción anafiláctica y los controles.
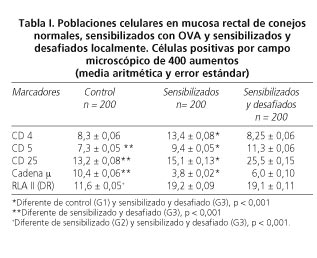
El marcador CD5 permite visualizar linfocitos T totales y determinadas células B
(Fig. 2) observándose, en nuestro modelo, un incremento en el grupo experimental.
Existe un marcado aumento en el número de células CD25+ activadas post desafío (receptor de interleuquina 2, IL-2) (Fig. 3).
Las células RLA II+ presentan un aspecto dendrítico y se distribuyen homogéneamente en toda la mucosa rectal (Fig. 4).
DISCUSIÓN
La ovoalbúmina es un antígeno soluble que administrado en forma subcutánea genera sensibilización con anticuerpos IgE específicos. El desafío posterior en individuos sensibilizados con el antígeno en cuestión, produce una respuesta alérgica anafiláctica en el órgano digestivo sometido a desafío.
Distintos autores han desarrollado modelos de alergia alimentaria con sensibilización y desafío, pero existen escasos trabajos que utilicen el recto como órgano de estudio. Este presenta como hechos destacables que la mucosa rectal pertenece al sistema inmune mucoso común y por lo tanto, los fenómenos inmunes de sensibilización generan una respuesta en los diferentes sitios efectores sometidos a desafío antigénico; en segundo término, la relativa accesibilidad del recto para realizar estudios diagnósticos, que posibiliten a partir de determinados patrones celulares e histopatológicos establecer diagnóstico certero (27).
Las alteraciones histopatológicas de tipo reacción anafiláctica en la mucosa rectal, se encuentran en su máxima expresión 4 h después del desafío. La edematización mucosa, la linfangiectasia y la infiltración de eosinófilos observados, expresan la acción de mediadores vasodilatadores y proinflamatorios. Este patrón es similar al hallado por otros autores y por nuestro grupo en trabajos anteriores, en intestino delgado y en otros sectores del tubo digestivo del conejo (11-28,29).
La ausencia de diferencias estadísticamente significativas entre los grupos estudiados para el marcador CD4, es un fenómeno diferente al ocurrido en enfermedades crónicas del intestino (enfermedad de Crohn), en la cual se produce un aumento marcado de células CD4+ en mucosa rectal (30,31). Esta diferencia podría corresponder a la característica del patrón histopatológico agudo de la reacción anafiláctica post desafío.
Es importante remarcar que, a diferencia de lo hallado en íleon de conejos sensibilizados y desafiados donde las células CD5+ se presentan tanto en lámina propia como a nivel intraepitelial (32) en recto no se observaron células CD5 positivas en el epitelio. Los linfocitos intraepiteliales del intestino delgado comprenden una población que reside en proximidades de la luz intestinal. Poseen una potente acción citolítica y capacidad inmunorregulatoria que les permiten mantener la integridad epitelial.
Es probable que esta diferencia entre íleon y recto se relacione con las distintas funciones que ambos órganos cumplen en el aparato digestivo, ya que el intestino delgado enfrenta una enorme cantidad de proteínas alimentarias potencialmente antigénicas. Por su parte, el recto no contacta macromoléculas alimentarias, pero existe una mayor concentración de microorganismos bacterianos de la flora saprofita en la región luminal.
El marcador CD25 se expresa en las células que activadas participan en la vertiente efectora de la respuesta inmune, durante los fenómenos de inflamación alérgica post-desafío. Si bien en nuestro trabajo no hemos estudiado el fenotipo de las células CD25+, numerosos autores coinciden en considerar a estas células macrófagos, por las características morfológicas que presentan. La mayor parte de las células son voluminosas y se encuentran fundamentalmente subyacentes al epitelio. En cuanto al tiempo de expresión del marcador CD25, nosotros lo determinamos 4 h post-desafío, coincidiendo con otros autores sobre la expresión génica que marca que la aparición del receptor de IL-2 se inicia 4 a 6 horas post-desafío (30,31).
La cadena µ corresponde a linfocitos B inmaduros que expresan en su superficie cadenas pesadas, reconocidas por anticuerpos específicos. Estas células sufren una marcada disminución en la mucosa rectal de los grupos experimentales. Esta disminución podría corresponder a la menor expresión de cadena µ en membrana de linfocitos B como consecuencia del cambio de isotipo (switching) o bien a una disminución real en el número de linfocitos B en el sitio estudiado a partir de una migración a otros sectores.
Es de interés remarcar que al igual que lo hallado por nosotros y otros autores (33), se observan en la mucosa, en proximidades del epitelio, células RLA-II positivas de aspecto dendrítico. En esta localización podrían corresponder a células dendríticas presentadoras de antígeno, si bien serían necesarios estudios de doble tinción para establecer el fenotipo. Otra característica de las células RLA-II+ es que en algunos sectores se encuentran como agregados subepiteliales dispersos.
Los resultados obtenidos nos permiten concluir que la sensibilización y el posterior desafío con OVA producen modificaciones cuantitativas de las células estudiadas. Estos datos cuantitativos pueden considerarse como patrones inmunopatológicos de la reacción anafiláctica aguda local en la mucosa rectal de animales sensibilizados por vía sistémica.
BIBLIOGRAFÍA
1. Brandtzaeg PE. Current understanding of gastrointestinal immunoregulation and its relation to food allergy. Ann N Y Acad Sci 2002; 964: 13-45. [ Links ]
2. Taylor SL, Hefle SL. Food science perspective on food allergy. Allergy 1998; 53: 5-7. [ Links ]
3. Holt PG. Mucosal immunity in relation to the development of oral tolerance/ sensitization. Allergy 1998; 53: 16-9. [ Links ]
4. Mayer L. Mucosal immunity and gastrointestinal antigen processing. J Pediatr Gastroenterol Nutr 2000; 30: 4-11. [ Links ]
5. Brandtzaeg P. The human intestinal immune system: Basic cellular and humoral mechanisms. Baillieres Clin Rheumatol 1996; 10: 1-24. [ Links ]
6. Strober W, James SP. The immunologic basis of inflammatory bowel disease. J Clin Immunol 1986; 6: 415-32. [ Links ]
7. van Ree R, Akkerdaas J, van Leeuwen A, Fernández Rivas M, Asero R, Knul-Bretlova A et al. New perspectives for the diagnosis of food allergy. Allergy Clin Immunol Int 1998; 12: 7-12. [ Links ]
8. Sampson HA. Food allergy. JAMA 1999; 278: 1888-94. [ Links ]
9. Placenti G, Bertolini A, Spezia E, Piscione T, Boner AL. Ability of new infants formula prepared from partially hydrolysed whey to induce anaphylactic sensitization: evaluation in guinea pig model. Allergy 1994; 49: 361-4. [ Links ]
10. Miller K , Laugee J, Meredith C. The monitoring of effects of food components on immunoreactivity in experimental animals. Allergy 1998; 53: 35-7. [ Links ]
11. Vinuesa M, Tanaka Y, Hakugawa J, Jae Bae S, Katayama I. In-situ expression of interleukin-4, 5 and 6 in Peyer's Patch from Ovalbumin (OVA)-sensitized BALB/c mice after oral challenge. Int Allergol 1997; 46: 243-7. [ Links ]
12. Sampson HA. Food allergy. Part I: Immunopathogenesis and clinical disorders. J Allergy Clin Immunol 1999; 5: 717-29. [ Links ]
13. Sabbah A, Drouet M, Luret MG, Giffard G, Minkhar M. Cross reactivity between cow's milk and goat's milk. Allerg Immunol 1997; 29: 203-14. [ Links ]
14. Ahmed T, Fuchs G. Gastrointestinal Allergy to food: A review. J Diarrheal Dis Res 1997; 15: 211-23. [ Links ]
15. Ramírez Cruz NE, Castrejón Vázquez MI, Espinoza Goldman MB, Martínez-Cairo Cueto S. Prevalence of latex-fruit syndrome in health workers with latex allergy. Rev Alerg Mex 2002; 49: 46-51. [ Links ]
16. Moller M, Kayma M, Vieluf D, Paschke A, Steinhart H. Determination and characterization of cross-reacting allergens in latex, avocado, banana, and kiwi fruit. Allergy 1998; 53: 289-96. [ Links ]
17. Mage RG. Immunology of lagomorphs. En: Pastoret P, Griebel P, Bazin H, Govaerts A, eds. Handbook of Vertebrate Immunology. San Diego: Academic Press, 1998. p. 223-60. [ Links ]
18. Fekete S, Bonori J. The effect of the fiber and protein level of the ration upon the cecotrophy of rabbit. J Appl Rabbit Res 1985; 8: 68-71. [ Links ]
19. Hollyster AG. Effects of water administrated probiotics and acidifiers on growth feed conversion and enteritis mortality of weaning rabbit. J Appl Rabbit Res 1989; 12: 143-7. [ Links ]
20. Fekete S. Recent finding and future perspectives of rabbit's digestive physiology. Cuni Sciences 1987; 4: 1-9. [ Links ]
21. Bassan N, Vinuesa M, Roma S, Pérez F. Biological model for detection of food antigens. Arch Latinoam Nutr 2002; 52: 249-56. [ Links ]
22. Cellier C, Cervoni JP, Leborgne M, Marteau P, Landi B, Cerf-Bensussan N, et al. Gluten-free diet induces regression of T-cell activation in the rectal mucosa of patients with celiac disease. Am J Gastroenterol 1998; 93: 1527-30. [ Links ]
23. Ensari A, Marsh M, Loft D, Morgan S, Moriarty K. Morphometric analysis of intestinal mucosa. V-Quantitative histologic and immunocytochemical studies of rectal mucosae in gluten-sensitivity. Gut 1993; 34: 1225-9. [ Links ]
24. Troncone R, Mazzarella G, Leone N, Mayer M, De Vincenzi M, Greco L, et al. Gliadin activates mucosal cell mediated immunity in cultured rectal mucosa from coeliac patients and a subset of their siblings. Gut 1998; 43: 484-9. [ Links ]
25. Kotani M. Generation and characterization of monoclonal antibodies against rabbit CD4, CD5 and CD11a antigens. J Imm Meths 1993; 157: 241-52. [ Links ]
26. Bassan N, Vinuesa M, Pérez F, Roma S, Bernardi S, Lagrutta M. Mastocitos Azul Alciano positivos en la mucosa del ciego de conejos normales y esplenectomizados, sensibilizados y desafiados con ovoalbúmina. Analecta Veterinaria 1998; 18: 15-20. [ Links ]
27. Honma K, Kohno Y, Hirano K, Shimojo N, Suzuki H, Hoshioka A, et al. Diagnosis of food allergy based on rectal mucosa cytology. Arerugi 1992; 41: 749-56. [ Links ]
28. Perdue M, Chung M, Gall G. Effect of intestinal anaphylaxis on gut function in the rat. Gastroenterol 1984; 86: 391-7. [ Links ]
29. Bassan N, Vinuesa M, Pérez F, Roma S, Bernardi S. Células enteroendocrinas intraepiteliales en ciego y apéndice de conejos sensibilizados con ovoalbúmina. Acta Gastroenterol Latinoam 1999; 29: 313-7. [ Links ]
30. Barois N, De Saint-Vis B, Lebecque S, Geuze HJ, Kleijmeer MJ. MHC Class II Compartments in human dendritic cells undergo profound structural changes upon activation. Traffic 2002; 3: 894-905. [ Links ]
31. Choy M, Walker-Smith J, Williams C, MacDonald T. Differential expression of CD25 (interleukin-2 receptor) on lamina propria T cells and macrophages in the intestinal lesions in Crohn's disease and ulcerative colitis. Gut 1990; 31: 1365-70. [ Links ]
32. Vinuesa M, Roma S, Bassan N, Pérez F. Characterization of intraepithelial T cell population in terminal ileum from ovalbumin (OVA) sensitized New Zealand rabbit. J All Clin Immunol 1997; 4: 74-6. [ Links ]
33. Kelsall B, Strober W. Distinct populations of dendritic cells are presents in the subepithelial dome and T cell regions of the murine Peyer's patch. J Exp Med 1996; 183: 237-47. [ Links ]

